4 ESO LOMCE: Biologia i Geologia
ÍNDEX 1. L'organització cel·lular dels éssers vius ►
2. Herència i genètica ►
3. La informació i manipulació genètica ►
3.1. Mutacions ►
3.2 Biotecnologia i enginyeria genètica ►
4. L'origen i l'evolució de la vida ►
5. Estructura de la Terra ►
6. Dinàmica de la Terra. Tectònica i relleu.►
7. La història de la Terra ►
8. Estructura i dinàmica dels ecosistemes ►
9. L'activitat humana i el medi ambient. ►

En el nivell molecular, els àtoms dels elements químics s'associen per formar molècules (aigua, proteïnes, etc,.). Diferents molècules s'associen per formar els orgànuls cel·lulars (cromosomes, ribosomes, cloroplasts, etc.).
Els orgànuls cel·lulars formen en conjunt una unitat superior: la cèl·lula, que realitza totes les funcions d'un éssers viu. La cèl·lula és la part més petita que té vida pròpia.
Molts éssers vius formats per una cèl·lula (unicel·lulars) i molts altres estan formats per moltes cèl·lules (pluricel·lulars). Els grups de cèl·lules que s'especialitzen en realitzar una mateixa funció formen un teixit (muscular, nerviós, conductor, etc.) diferents teixits s'agrupen i formen els òrgans (ronyó, cor, etc.). Els òrgans s'agrupen i formen els aparells i sistemes (digestiu, nerviós, etc.). Els diferents sistemes s'agrupen i funcionen coordinadament donant lloc a un ésser pluricel·lular.
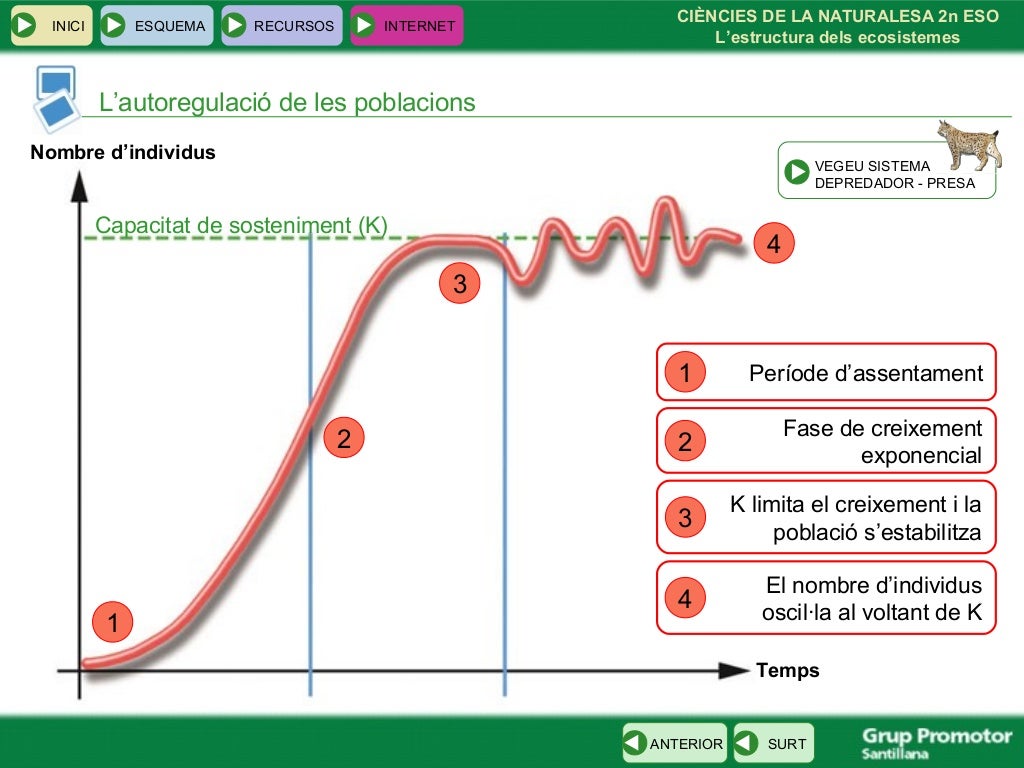
Els éssers vius formen poblacions. Una població és una agrupació d'individus de la mateixa espècie que viuen en una zona determinada, i que difícilment poden viure aïllats ja que necessiten relacionar-se entre ells per la reproducció, l'alimentació, etc. L'associació de poblacions de distintes espècies que tenen un dinàmica pròpia constitueixen una comunitat (vegetació d'un bosc, animals d'un llac, etc.).
Les poblacions estan molt lligades al medi on viuen, i en conjunt formen l'ecosistema, que és el conjunt d'éssers vius que viuen en un lloc, les relacions que estableixen entre ells i el medi.
El conjunt d'ecosistemes forma la biosfera, zona de la Terra on existeix la vida.
1. Els nivells d'organització
Els nivells d'organització són els diferents graus de complexitat en què s'organitza la matèria viva.Els elements de cada nivell s'agrupen per formar altres nivells més complexos, amb característiques i propietats diferents que els elements del nivell anterior.
Per facilitar l'estudi de l'estructura (anatomia) i funcionament (fisiologia) dels organismes vius, es defineixen diferents nivells d'organització.
Abiòtic
Nivell atòmic: àtoms. L'àtom és la part més petita de la matèria que manté les seves propietats.
Nivell molecular: una molècula és el resultat de la unió de diferents àtoms.
Biomolècules: molècules que formen els éssers vius. Poden ser:
Biomolècules Inorgàniques: les podem trobar en la matèria viva i en la inerta. Pe: aigua i sals minerals.
- Hi tenen lloc totes les reaccions químiques.
- Formen estructures (closques, esquelet,...)
- Glúcids o hidrats de carboni
- S'utilitzen per emmagatzemar energia (midó - plantes, glucògen - animals)
- Forma les membranes cel·lulars (fosfolípids o colesterol)
- La unió dels aminoàcids forma les proteïnes.
- Funció defensiva: anticossos
- La unió de nucleòtids forma els àcids nucleics. N'hi ha de dos tipus:
- ARN (àcid ribonucleic): participa en la síntesi de proteïnes.
Biòtic
La cèl·lula és la unitat funcional i estructural dels organismes vius capaç de reproduir-se (Teoria cel·lular).
Nivell de teixit: conjunt de cèl·lules amb una mateixa funció i un mateix origen. Pe: teixit muscular, epitalial, ossi,...
Nivell d'òrgan: els òrgans estan formats per diferents teixits que s'agrupen per dur a terme una funció. Pe: estómac, cor, ronyó,...
Nivell sistema: conjunt d'òrgans semblants que realitzen una funció que estan formats per un mateix tipus de teixit. Pe: sistema esquelètic, muscular, nerviós,...
Nivell aparell: conjunt d'òrgans diferents, cada un amb una funció i que participen en una o diverses funcions. Pe: aparell digestiu , locomotor,...
Nivell d'organisme: ésser viu.
Nivell de població: conjunt d'individus de la mateeixa espècie.
Nivell de comunitat: conjunt de poblacions que comparteixen un mateix espai.
Nivell d'ecosistema: és el conjunt d'èssers vius (biocenosi) que habiten en un lloc (biòtop), les relacions que s'estableixen entre aquests organismes, a més de les característiques del lloc i les relacions entre el medi i els organismes.
En els organismes unicel·lulars el nivell cel·lular i el nivell organisme coincideixen.


2. La cèl·lula unitat de vida
Leeuwenhoek va recollir els fruits de la intel·ligent col·locació de les lents convergents, ideada pels germans Jansen (1590), i va saber construir el primer microscopi, realitzant, a més, encertades observacions microscòpiques.






Gràcies a aquestes millores instrumentals i tècniques, en 1824 Dutrochet va formular per primera vegada en la història de la Biologia que tots els animals i plantes estaven formats per cèl·lules de diferents tipus.
Van ser els científics alemans M.J. Schleiden (botànic) i T.Schawann (zoòleg) qui al 1838 i 1839 van elaborar la teoria cel·lular.
Cada cèl·lula és una unitat constitucional i funcional dels éssers vius, capaç de mantenir la seva existència de manera independent.
Al 1858, Rudolf Virchow va publicar el seu famós tractat de Patologia cel·lular, en el qual, entre d’altres coses, no només defensava la feina de Schleiden i Schawnn, sinó que la completava amb la seva famosa frase omnis cellula ex cellula (tota cèl·lula provés d’una altra cèl·lula).
Així la teoria cel·lular quedava definida per aquests tres principis:
- Cada cèl·lula és capaç de mantenir la seva pròpia vitalitat per ella tota sola.
Va ser Santiago Ramón y Cajal qui va demostrar la individualitat neuronal demostrant i donant validesa universal a la teoria cel·lular.
Santiago Ramon i Cajal, metge, històleg i professor universitari espanyol guardonat amb el Premi Nobel de Medicina o Fisiologia el 1906, descobreix, l’any 1888, durant la seva estada a Barcelona, aplicant el mètode de tinció de Camillo Golgi, els mecanismes que governen la morfologia i els processos connectius de les cèl·lules nervioses, de la matèria grisa i del sistema nerviós cerebroespinal. La seva teoria va ser acceptada el 1889, durant el Congrés de la Societat Anatòmica Alemanya celebrat a Berlín. El seu esquema estructural del sistema nerviós com un aglomerat d’unitats independents i definides va passar a conèixer-se com “doctrina de la neurona” i en ella destaca la llei de la polarització dinàmica, model capaç d’explicar la transmissió unidireccional de l’impuls nerviós.
L’any 1906 fou guardonat, juntament amb l’italià Camillo Golgi, amb el Premi Nobel de Medicina o Fisiologia “pels seus estudis sobre el sistema nerviós”.


2.1. Teoria cel·lular
- La cèl·lula és la unitat estructural dels éssers vius. Tots els éssers vius estan formats per una o més cèl·lules.
- La cèl·lula és la unitat funcional dels éssers vius. És la unitat mínima capaç de dur a terme les funcions vitals d'un ésser viu. (Schleiden, Schwann - 1838)
- Tota cèl·lula procedeix, per divisió, d'una altra cèl·lula prexistent. (Virchow - 1855)
3. Tipus de cèl·lules: procariotes
- Definició de - Procariotes
Procariotes: cèl·lules que tenen el material genètic dispers en el citoplasma formant el nucleoide (del grec pro - abans i karyon - nucli). Cèl·lules sense embolcall nuclear.
- Parts: estructura i funció
a - Nucleoide
b - Citoplasma
c - Ribososmes (70S)
d - Membrana plasmàtica
e - Pared cel·lular: Gram + i Gram -
f - Flagels, cilis, fímbries
g - Càpsula
h - Bacteris: formes, importància
![]() Organismes procariotes
Organismes procariotes
4. Tipus de cèl·lules: eucariotes
4.1. Definició de - Eucariotes.
Cèl·lula eucariota (del grec eu,'veritable', i karyon, 'nou' o 'nucli') és aquella cèl·lula que té el material genètic delimitat per una membra lipídica o embolcall nuclear format el nucli cel·lular.

4.2. Origen de la cèl·lula eucariota. Teoria endosimbiòntica.
 Sovint es diu que els mitocondris són les centrals energètiques de les cèl·lules eucariotes, ja que aquests orgànuls s'encarreguen de obtenir energia mitjançant la respiració cel·lular, és a dir, oxiden matèria orgànica procedent de la digestió per obtenir energia que emmagatzemen en molècules d'ATP.
Sovint es diu que els mitocondris són les centrals energètiques de les cèl·lules eucariotes, ja que aquests orgànuls s'encarreguen de obtenir energia mitjançant la respiració cel·lular, és a dir, oxiden matèria orgànica procedent de la digestió per obtenir energia que emmagatzemen en molècules d'ATP. La teoria endosimbiòtica proposada per Lynn Margulis el 1971 explica l'origen de les cèl·lules eucariotes a partir d'una primitives cèl·lula procariota (anomenada eucariota), capaç de fagocitar bacteris sense digerir-los de manera que quedaven assimilats i es convertien en els actuals mitocondris. Simultàniament, les cèl·lules van augmentar la seva complexitat, i van donar lloc a un sistema intern de membranes que van permetre l'aparició d'orgànuls i de l'embolcall nuclear. En aquesta relació, l'organisme procariota fagocitat aconseguia matèria orgànica que en reaccionar amb l'oxigen produïa energia que emmagatzemaven en les molècules d'ATP, part de les quals passaven a la cèl·lula hoste. Aquest benefici mutu va fer que la cèl·lula fagocitada arribés a forma part de l'organisme hoste, fins a convertir-se en part d'ella: el mitocondri.
Altres factors que recolzen aquesta teoria sobre l'origen bacterià dels mitocondris són la seva semblança amb aquestes cèl·lules procariotes, ja que presenten una forma i grandària molt similars, una estructura de doble membrana, un ADN circular, ribosomes de procariota, components semblants en la seva membrana i la forma en que produeix energia.
Per tant, a les nostres cèl·lules, a més de l'ADN del nucli també tenim ADN mitocondrial.
L'ADN mitocondrial s'hereta només per via materna, ja que quan un espermatozoide fecunda un òvul només hi penetra el nucli, i em queda fora la resta de la seva matèria cel·lular, per això en el zigot tots el mitocondris existents provenen de l'òvul. Una altra característica important que cal destacar de l'ADN mitocondrial és que gairebé no es modifica. Els únics canvis possibles amb ADN mitocondrial són deguts exclusivament a les mutacions. Segons els càlculs estadístics que s'han realitzat fins ara, s'informa que en l'èsser humà, cada10.000 anys aproximadament, es produeix una mutació en un dels nucleòtids de l'ADN mitocondrial. Fent operacions es pot concloure que entre una dona que hagués nascut fa 400.000 anys i un descendent directe actual hi hauria uns 4 nucleòtids diferents.
Coneixent el nombre de mutacions s'ha pogut calcular que la dona de la nostra espècie, anomenada Eva mitocondrial, de la qual provenen els nostres mitocondris, devia haver viscut a Àfrica en una època compresa entre els 140.000 i els 290.000 anys d'antiguitat.

Migracions humanes en tot el món segons les dades de l'ADN mitocondrial.
4.3. Parts: estructura i funció.
a - Membrana cel·lular
b - Paret cel·lular
c - Nucli
d - Citoplasma
e - Orgànuls cel·lulars
f - Orgànuls membranosos
g - Orgànuls no membranosos
a - Membrana cel·lular
Model de Mosaic fluid (Singer i Nicholson - 1972)
- Doble capa de lípids associada a molècules proteïques.
- Les proteïnes poden estar només en la superfície d'una de les cares o ocupar les dues capes.
- Totes les molècules poden intercanviar la seva posició en la bicapa amb altres molècules.
Segons aquest model la membrana presenta dues propietats:
- Estructura dinàmica: els lípids es mouen i estableixen enllaços amb lípids del seu voltant. L'autoacoblament permet l'autoreparació de la mb i els processos d'edocitosi i exocitosi.
- Estructura asimètrica: és asimètrica en la cara ext-int. En l'exterior el glucocàlix té funcions de reconeixement cel·lular (per a la fecundació, per l'immunitat, reconeixemnt de cel del mateix teixit o reconeixment de virus de la cel hoste)
Funcions:
- És impermeable per a les substàncies polars i permeable a les apolars.
- Processos d'endocitosi i exocitosi per autoacoblament.
- Reconeixement cel·lular.
- Ancoratge exterior - matriu i intern - citoesquelet
- Unions intercel·lulars.
- Transducció de senyals.
- Activitat enzimàtica.


- Xarxa de fibres de cel·lulosa (polisacàrid estructural (ß-D- glucoses amb enllaç ß 1-4))
- Matriu:
- Altres:
- Suberina: protegeix i aïlla del foc (escorça)
- Cutina: impermeabilitza (epidermis de la fulla)
- Carbonat de calci i sílice: rigidesa (epidermis de la fulla)
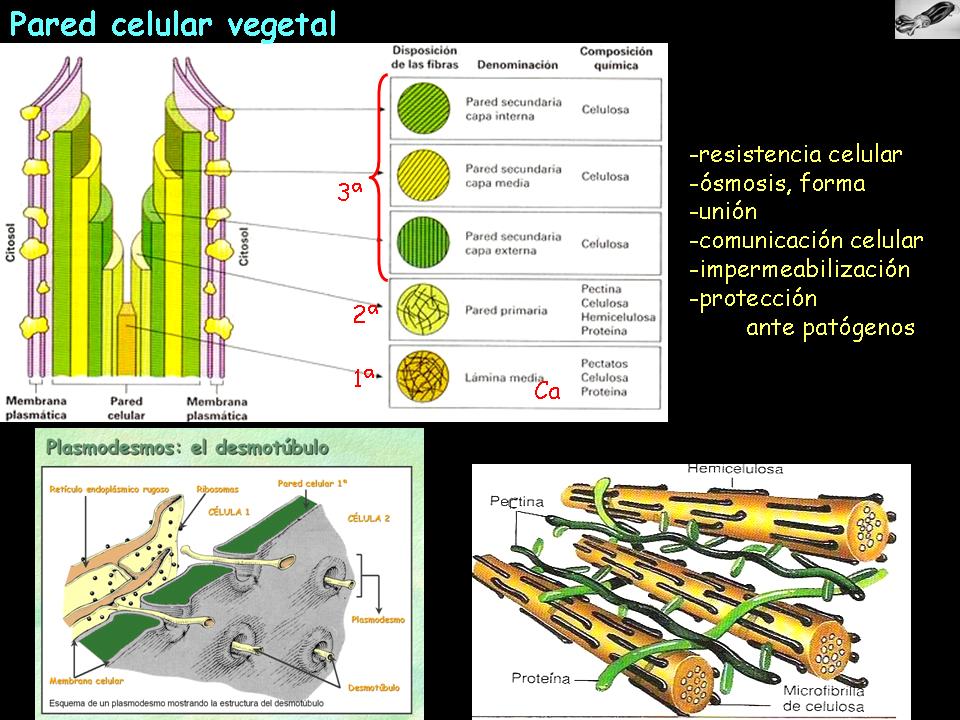
- Evita la ruptura de la cèl·lula per pressió osmòtica
c - Nucli
- Nucleoplasma: solució aquosa que es troba a l'interior del nucli.
- ADN - àcid desoxiribonucleic - material genètic: conté la informació i controla les funcions cel·lulars.
L'ADN està associat a proteïnes i forma la cromatina.
Quan la cèl·lula s'ha de dividir, la cromatina es condensa i s'enrotlla en espiral formant unes estructures gruixades anomenades cromosomes (L'èssers humà té 46 cromosomes). L'ADN dels cromosomes es reparteix entre les cèl·lules filles.
Nucleòl: quan la cèl·lula està realitzant les funcions vitals (viu) necessita la informació que conté l'ADN, per tant s'han de fer còpies d'aquesta informació. El nucleòl és una zona densa del ADN que es veu al microscopi i és on s'acumula l'ADN que s'està copiant, les proteïnes que el copien i l'ARN que és la còpia.

c- Citoplasma
Solució aquosa que conté:
- substàncies dissoltes
- fibres de proteïnes formant el citoesquelet
- orgànuls cel·lulars
d - Orgànuls cel·lulars
Estructures que duen a terme diferents funcions.
La presència d'uns orgànuls o altres i la seva abundància depèn de la funció que realitza cada cèl·lula.
e - Orgànuls membranosos
- Mitocondris
Ovalats
Tenen dues membranes. L'externa és llisa i l'interna amb nombrosos plecs, anomenats cretes. Entre ells es distingeix l'espai intermembranal.
En la membrana interna es troben les proteïnes que permeten fer la respiració cel·lular.
En ell hi podem trobar molules d'ADN mitocondrial circular i de doble filament, com les bacterianes.
I ribosomes semblants als bacterians.
- Funció:
S'hi du a terme la respiració cel·lular. (Glucosa + oxígen —►Diòxid de carboni + aigua + Energia - ATP)


- Cloroplasts
Orgànul format epr dues membranes: membrana externa, membrana interna i espai intermembranal.
L'estroma és el medi intern que conté: l'ADN circular de doble hèlix, com els dels bacteris, ribosomes 70S, inclusions de midó i lipíques i altres enzims.
Dins l'estroma trobam el tilacoides o lamel·les que són sàculs aplanats. Presenten una membrana tilacoïdal i una cavitat interior que rep el nom de lumen o espai tilacoïdal. A les membranes dels tilacoides s'ubiquen els pigments fotosintètics i les proteïnes que fan possible el procés de fotosíntesi. El tilacoides poden ser d'estroma si són allargats per l'estroma o de grànuls si són menuts amb forma de disc i apilats.
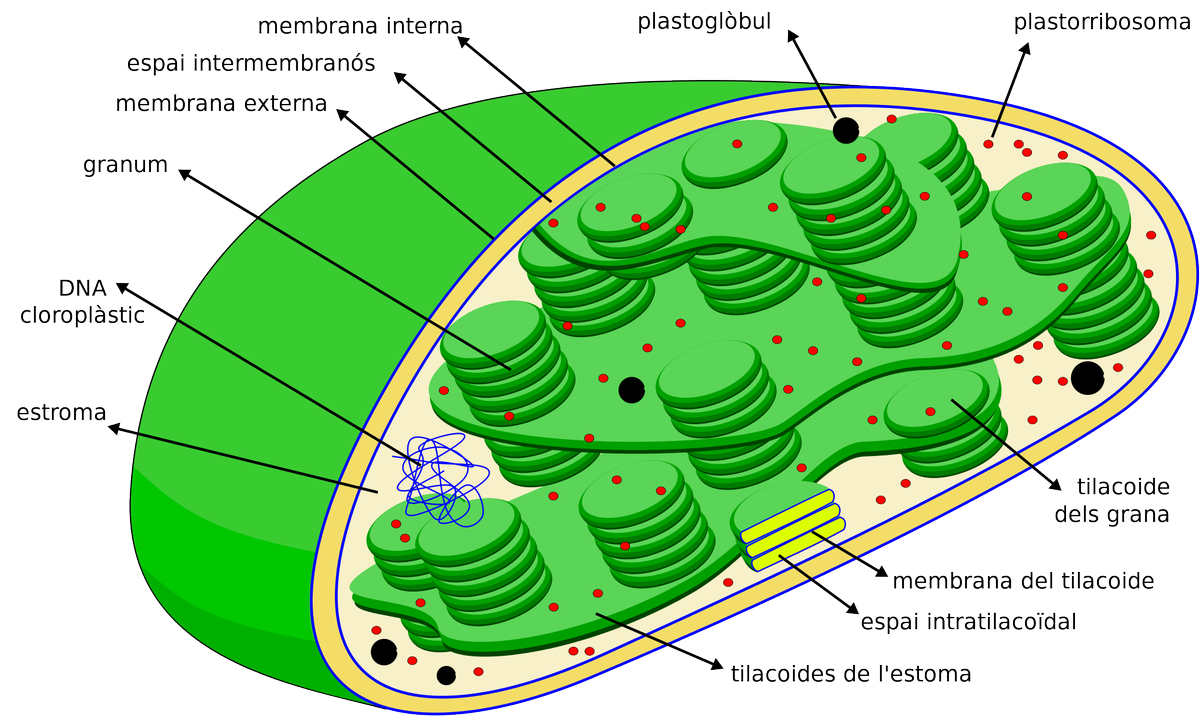
Són els orgànuls encarregats de fer la fotosíntesi.
Presenta dues fases:
1. Fase lluminosa (depenents de la llum): té lloc dins el tilacoide i genera ATP i NADPH (poder reductor) per poder utilitzar-ho en la següent fase.
2. Fase obscura (independent de la llum) (Cicle de Calvin): té lloc en l'estroma. Gràcies al ATP i al NADPH fabricats en l'anterior fase es fixa CO2 i es genera matèria orgànica.

- Reticle endoplasmàtic
Reticle endoplasmàtic rugós - RER
- Estructura: format per un conjunt de sacs aplanats comunicats entre ells i que s'expandeixen pel citoplasma.
Es situen al voltant del nucli.
A la cara externa hi té adherits els ribosomes.
- Funció:
Emmagatzema i transporta les proteïnes que sintetitzen els seus ribosomes.


Reticle endoplasmàtic llis - REL
- Estructura: format per un conjunt de tubs aplanats comunicats entre ells i que s'expandeixen pel citoplasma.
La seva membrana no té ribosomes.
- Funció:

- Aparell de Golgi
- Estructura: són una sèrie de sacs aplanats i en forma de disc superposats i no comunicats. En la seva perifèria s'emeten vesícules.
L'aparell de Golgi és un orgànul format per un o més disctiososmes. Un dictiosoma és una agrupació en paral·lel d'entre 4 i 8 sàculs o cisternes. Al dictiosoma s'hi distingeixen dues cares: - cara cis o de formació (cara pròxima al RER) i - cara trans o de maduració orientada cap a la membrana plasmàtica.
- Funció:
Transport, maduració, acumulació i secreció de proteïnes procedents del RE.
Síntesi de polisacàrids - glúcids.
Rep i modifica les substàncies procedents del reticle endoplasmàtic (RER - proteïnes, REL - lípids) hi afegeix glúcids..
Allibera totes aquestes substàncies mitjançant petites vesícules.
- Lisosomes
- Estructura:
Conté hidrolasas àcides que digereixen substàncies de reserva o de rebuig.
Les vesícules que digereixen els productes cel·lulars (digestió: molècules complexes per obtenir molècules senzills) s'anomenen lisosomes.

- Vacúols
Orgànuls membranosos (1 membrana).
Funció de transport entre orgànuls i entre aquest i l'exterior i d'emmgatzematge d'aigua o substàncies específiques (reserva energètica, productes de rebuig, funció esquelètica).
Intervenen en la regulació de la quantitat d'aigua i sals de la cèl·lula.
En les cèl·lules animals, els vacúols solen ser menuts. En les cèl·lules vegetals solen ser molt grans i, en general, només n'hi ha un o dos en cada cèl·lula.
- Peroxisomes
Orgànuls membranosos (1 membrana).
- Funció:
Conté la catalasa, que és un enzim que transforma el peròxid d'hidrogen (molt tòxic) en aigua i oxigen.
Tenen una funció detoxificant, degraden susbtàncies tòxiques.
f - Orgànuls no membranosos
- Ribosomes
 - Estructura:
- Estructura:
Són partícules no membranoses formades per ARN (àcid ribonucleic) i proteïnes.
Formades per dues subunitats.
Poden estar dispersos en el citoplasma o adherits a les membranes del RER.
- Funció:
Sintetitzar molècules de proteïnes.
Procariotes 70S. Eucariotes 80S.
- Centrosoma
- Estructura:
Són dos cilindres perpendiculars (centríols) formats per túbuls de proteïnes.
Es troba al voltant del nucli.
- Funció:
Intervé en la divisió cel·lular, regulant el repartiment de cromosomes a les cèl·lules filles.
Forma part dels cilis i flagels que mouen algunes cèl·lules (espermatozoides) o ajuden a moure el medi extern (cèl·lules de la tràquea).
4.4. Cèl·lula animal vs vegetal.

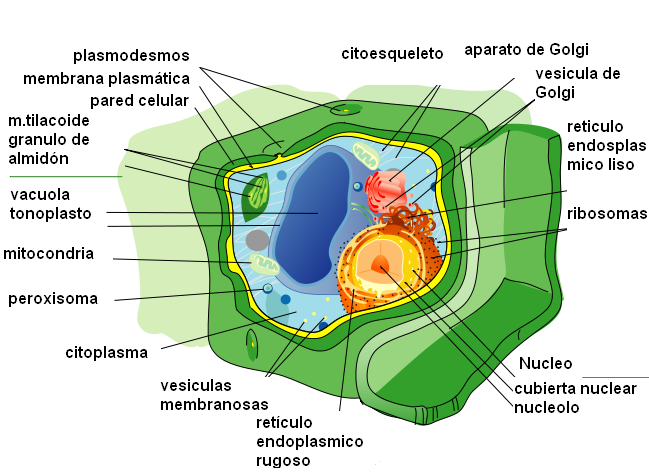
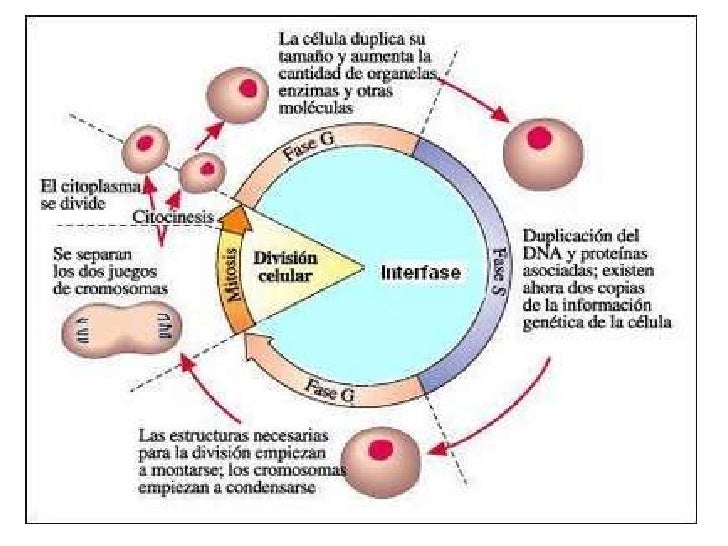
5. Cicle cel·lular.
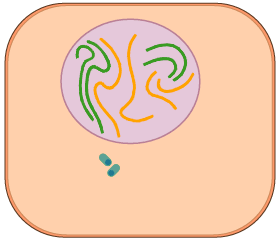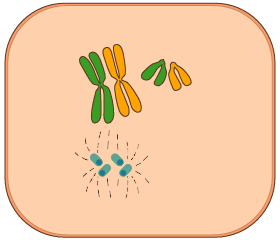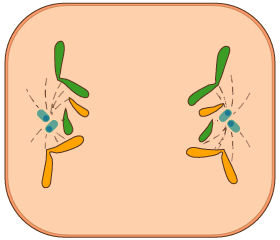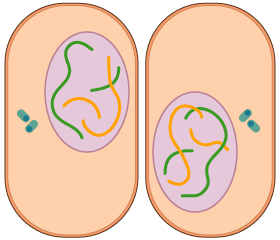
La interfase és un període de gran activitat cel·lular. La interfase està composta per fases G1, S i G2, i normalment representa el 90% del total del cicle cel·lular.
L'altra fase concreta del cicle cel·lular és la de divisió cel·lular, fàcilment observable al microscopi, i que inclou una divisió nuclear (mitosi) i una divisió citoplasmàtica (citocinesi). Tota la fase de divisió cel·lular rep el nom de fase M (M = mitòtica).
- G1 - G0. (G = gap, en anglès separació) És una fase de creixement que s'inicia immediatament després de la divisió cel·lular. És una etapa d'intensa activitat metabòlica on els gens es transcriuen i es tradueixen per sintetitzar les proteïnes necessàries per al creixement cel·lular. Acostuma a durar aproximadament 11 hores.
Si la fase G1 s'allarga molt en el temps es diu que la cèl·lula ha entrat en G0; cosa que passa amb les cèl·lules madures o que ja s'han diferenciat (cèl·lules nervioses, cel musculars cardíaques). Entren en un estadi de quietud pel que fa a la divisió encara que segueixen realitzan totes les funcions que li han estat encomanades. En algunes cèl·lules molt especialitzades la diferenciació cel·lular no permet que es tornin a dividir; en altres cèl·lules poden passar de G0 a G1 i dividir-se en ser correctament estimulades.
- S. Quan la cèl·lula assoleix unes determinades dimensions, s'ha de preparar per a la seva divisió; per això duplica prèviament el seu contingut genètic amb la finalitat que cada cèl·lula filla contingui una còpia idèntica del genoma (conjunt d'ADN de la cèl·lula). Aquesta és una fase de síntesi i replicació de l'ADN i finalitza quan el contingut d'ADN del nucli s'ha duplicat. Té una durada aproximada de 8 hores.
- Mitosi o divisió cel·lular. És l'etapa final del cicle, quan les cèl·lules es divideixen i reparteixen equitativament el seu contingut nuclear (mitosi) i citoplasmàtic (citocinesi) entre les dues cèl·lules filla. En primer lloc la cromatina que ha estat duplicada en la fase S es condensa i forma unes estructures - cromosomes- que han d'ésser alineats separats i desplaçats cap als extrems de la cèl·lula. En segon lloc, el citoplasma s'ha de segmentar de manera que asseguri que cada cèl·lula filla obtingui no solament un conjunt complet de cromosomes, sinó també els elements i orgànuls citoplasmàtics.


6. Els cromosomes
Cromosoma (del grec chroma, "color" i soma, "cos" o "element") és cadascun dels petits cossos en forma de bastonets què s'organitzen a la cromatina del nucli cel·lular durant les divisions cel·lulars (mitosi i meiosi) dels organismes eucariotes.
La cromatina és un material microscòpic que porta la informació genètica dels organismes eucariotes i està composta d'ADN associat a proteïnes especials anomenades histones. Aquest material es troba al nucli de les cèl·lules eucariotes i té l'aspecte d'un embull de fils prims. Quan el nucli cel·lular comença el procés de divisió, aquest embull de fils inicia un fenomen de condensació progressiu que finalitza en la formació d'entitats diferenciades i independents, els cromosomes. Per tant, cromatina i cromosoma són dos aspectes morfològicament diferents d'una mateixa cosa.

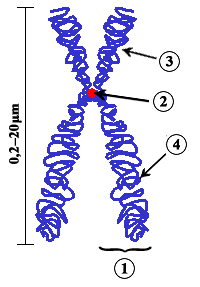 1) Cromàtide, cadascuna de les parts idèntiques d'un cromosoma després de la duplicació de l'ADN.
1) Cromàtide, cadascuna de les parts idèntiques d'un cromosoma després de la duplicació de l'ADN. 2) Centròmer, el lloc del cromosoma en el qual ambdues cromàtides es toquen.
3) Braç curt.
4) Braç llarg.
5) Telòmer
- Nombre de cromosomes: 46cr
Cèl·lules diploides 2n: dues dotacions cromosòmiques (cromosomes homòlegs).
Cèl·lules haploides n: una dotació cromosòmica (no tenen cromosomes homòlegs)
Els eucariotes de reproducció asexual tenen una quantitat n de cromosomes, mentre que els eucariotes de reproducció sexual tenen 2n cromosomes (tenen dues còpies de cada cromosoma), excepte els gàmetes que també en tenen només n. Quan s'examina la longitud d'aquests cromosomes i la situació del centròmer sorgeix el segon tret general: per cada cromosoma amb una longitud i una posició del centròmer determinades existeix un altre cromosoma amb trets idèntics, és a dir, gairebé tots els cromosomes es troben formant parelles. Els membres de cada parella s'anomenen cromosomes homòlegs.
- Segons la posició del centròmer
- Submetacèntrics
- Telocèntrics
- Cromosomes sexuals o heterocromosomes
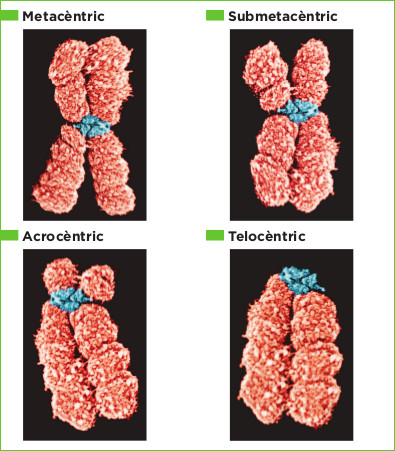

A l'àmbit clínic, el concepte de cariotip s'usa amb freqüència per referir-se a un cariograma, el qual és un esquema, foto o dibuix dels cromosomes d'una cèl·lula metafàsica ordenats d'acord a la seva morfologia (metacèntrics, submetacèntrics, acrocèntrics i telocèntrics) i mida, que estan caracteritzats i representen a tots els individus d'una espècie.
El cariotip és característic de cada espècie, igual que el nombre de cromosomes, l'ésser humà té 46 cromosomes (23 parells perquè som diploides o 2n) en el nucli de cada cèl·lula, organitzats en 22 parells autosòmics i 1 parell sexual (home XY i dona XX).

7. Divisió cel·lular
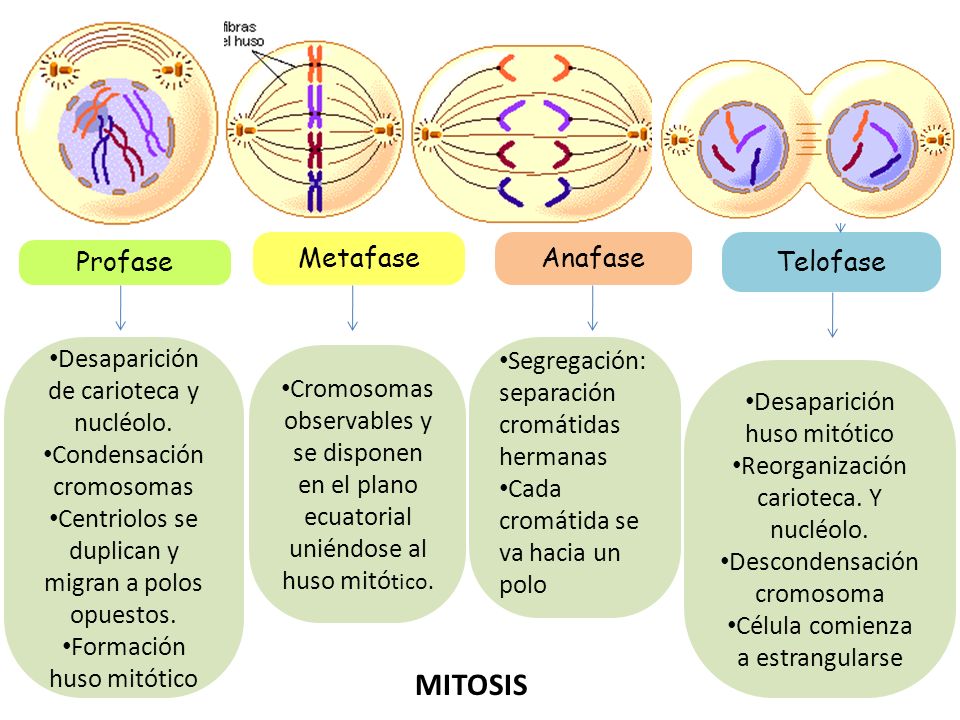
- Mitosi
- Profase
- Metafase
- Anafase
- Telofase
- Citocinesi
És una fase del cicle cel·lular en què és produeix la divisió del nucli. Procés de divisió de les cèl·lules somàtiques dels eucariotes, que consisteix, fonamentalment, en una divisió longitudinal dels cromosomes i en una divisió del citoplasma.
Aquest procés dóna com a resultat la constitució de dues cèl·lules filles.
La funció de la mitosi és assegurar el repartiment ordenat dels cromosomes a les cèl·lules filles, la dotació de les quals ha d’ésser idèntica a la de la cèl·lula mare.
En la mitosi podem distingir la profase, la metafase, l’anafase i la telofase.
- Durant la metafase cada centròmer de cada cromosoma s’uneix a una fibra del fus, i els cromosomes, cadascun dels quals és compost de dues cromàtides, s’estenen a la part equatorial de la cèl·lula.
- A la telofase una membrana nuclear envolta cadascun dels dos agrupaments de cromosomes i es trenquen les fibres del fus. Un cop dividit i reconstituït el nucli original en dos de nous, amb els nuclèols respectius, es produeix la divisió del citoplasma o citocinesi; això dóna com a resultat la constitució de dues cèl·lules filles.
8. Meiosi.
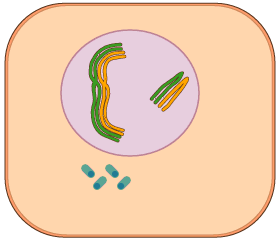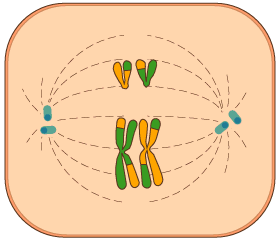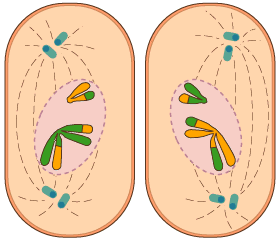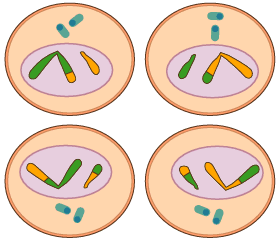 La meiosi és un procés especial de divisió cel·lular mitjançant el qual s'originen cèl·lules amb la meitat de cromosomes que la cèl·lula mare.
La meiosi és un procés especial de divisió cel·lular mitjançant el qual s'originen cèl·lules amb la meitat de cromosomes que la cèl·lula mare. La meiosi només es produeix en les cèl·lules sexuals o gàmetes.
La meiosi consta de dues divisions consecutives denominades primera divisió meiòtica i segona divisió meiòtica.
La primera divisió és reduccional (ja que les cèl·lules queden amb la meitat de cromosomes que la cèl·lula mare), mentre que la segona divisió meiòtica és pràcticament una mitosi normal.
Abans de la meiosi, igual que passa en la mitosi, es duplica l'ADN de la cèl·lula. Així, si partim d'una cèl·lula amb 2n cromosomes, després de les dues divisions meiòtiques tindrem quatre cèl·lules amb n cromosomes cadascuna.
- Metafase II els cromosomes es disposen en la placa equatorial, de forma que el pla equatorial talla els centròmers de cada cromosoma.
- Telofase II i la citocinesi s'aconsegueixen quatre cèl·lules, cadascuna d'elles amb la meitat de cromosomes que la cèl·lula mare original i una composició química única i diferent de la de les altres cèl·lules.

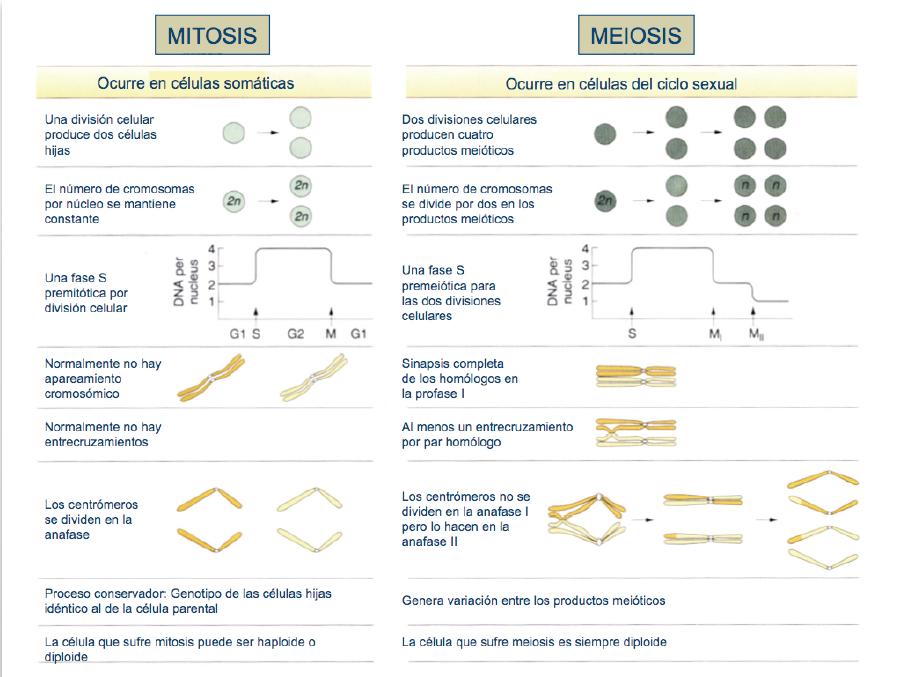
9. Mitosi vs Meiosi.


10. Gametogènesi.
- Espermatogènesi: És el procés de formació de gàmetes masculins (espermatozoides). Té lloc als testicles. Cada cèl·lula mare diploide produeix quatre cèl·lules haploides, que per un procés de diferenciació cel·lular donen lloc a quatre espermatozoides. Dos d'aquests espermatozoides porten un cromosoma X i els altres dos, un cromosoma Y.
- Oogènesi: És el procés de formació de gàmetes femenins (òvuls). Té lloc als ovaris. Cada cèl·lula mare diploide produeix quatre cèl·lules haploides, un òvul, que rep gairebé tot el citoplasma, i tres corpuscles polars, que no tenen cap utilitat i degeneren. Tots els òvuls porten un cromosoma X.

11. Càncer
Definició
La paraula càncer deriva del llatí, i aquesta del grec karkinos, i significa 'cranc'.
Es diu que les formes corrents de càncer avançat adopten una forma abigarrada, amb ramificacions, que s’adhereix a tot el que agafa, amb l’obstinació i forma semblant a la d’un cranc marí, i d’aquí deriva el seu nom.
El terme oncologia prové del grec onkos, tumor. És la part de la medicina que estudia els tumors o neoplàsies, sobretot malignes.

Història
El que ha passat a la història com el papir d'Edwin Smith demostra que els egipcis tenien un coneixement bastant exacte d’òrgans humans com ara el cor, el fetge, la melsa, els ronyons i els urèters, i la vesícula, a més de tractar amb molta més racionalitat de la qual se’ls suposava certs procediments quirúrgics. Però hi ha un fet que resulta més sorprenent encara i és que en el papir apareix la primera descripció escrita d’un càncer. En aquest es descriuen vuit casos de càncer de mama, que són tractats amb cauterització, encara que l’escrit diu de la malaltia que "no tenia tractament".
Nomenclatura oncològica
1. Les cèl·lules neoplàsiques - tumorals proliferants, és a dir, les cèl·lules que formen el tumor pròpiament dit.
2. El seu estroma de sosteniment, constituït per teixit connectiu i vasos sanguinis. Es tracta de teixits no tumorals la formació del qual ha estat induïda pel mateix tumor.
S’usen dos criteris de classificació: en primer lloc el seu caràcter benigne o maligne, i en segon lloc el teixit en què es formen.
Segons el comportament dels tumors:
1. Tumors benignes: El seu nom acaba en el sufix-oma, simplement, i segons l'origen del teixit del qual procedeixin els tumors benignes, poden ser: fibroma (teixit conjuntiu fibrós), mixoma (teixit conjuntiu lax), lipoma (teixit adipós), condroma (teixit cartilaginós), osteoma (teixit ossi), hemangioma (vasos sanguinis), limfangioma (vasos limfàtics), meningioma (meninges), tumor glómico (teixit nerviós de sustentació), leiomiomes (teixit muscular llis), rabdomioma (teixit muscular estriat ), papiloma (teixit epitelial formant papil), adenoma (teixit glandular), teratoma (cèl·lules totipotencials), nevus (melanòcits)
Alguns dels tumors benignes derivats de teixit epitelial acaben amb el sufix "adenoma, si bé hem de tenir en compte que existeixen múltiples excepcions a les normes de nomenclatura tumoral. Per exemple: El tumor benigne de melanòcits es denomina Nevus o nevo, i la seva forma maligna, Melanoma.
2. Tumors malignes o càncer:
Es considera a vegades sinònim dels termes 'neoplàsia' i 'tumor', però, el càncer sempre és una neoplàsia o tumor maligne.
El càncer, es pot originar a partir de qualsevol tipus de cèl·lula en qualsevol teixit corporal, no és una malaltia única sinó un conjunt de malalties que es classifiquen en funció del teixit i cèl·lula d’origen.
Existeixen diversos centenars de formes diferents, essent tres els principals subtipus:
- Els sarcomes procedeixen del teixit connectiu com ossos, cartílags, nervis, vasos sanguinis, músculs i teixit adipós.
- Els carcinomes procedeixen de teixits epitelials com la pell o els epitelis que entapissen les cavitats i òrgans corporals, i dels teixits glandulars de la mama i pròstata. Els carcinomes inclouen alguns dels càncers més freqüents. Els carcinomes d’estructura similar a la pell es denominen carcinomes de cèl·lules escamoses. Els que tenen una estructura glandular es denominen adenocarcinomes.
- En el tercer subtipus es troben les leucèmies i els limfomes, que inclouen els càncers dels teixits formadors de les cèl·lules sanguínies. Produeixen inflamació dels ganglis limfàtics, invasió de la melsa i moll d’os, i sobreproducció de cèl·lules blanques immadures.
Epidemiologia
El càncer és la segona causa principal de mort, darrere de les malalties cardíaques. No obstant això, les morts per malalties cardiovasculars estan disminuint, mentre que les morts per càncer estan augmentant. S’estima que al llarg del segle XXI, el càncer és la principal causa de mort en els països desenvolupats. Malgrat això, s'ha produït un augment en la supervivència dels pacients amb càncer.
Causa del càncer
És desconeguda, però es coneixen molts factors de risc que ho precipiten.
El principal factor de risc és l’edat o l’envelliment, ja que dues terceres parts de tots els càncers ocorren a qualsevol edat.
El segon factor de risc és el tabaquisme, i el segueixen la dieta, el sedentarisme, l’exposició solar i altres estils de vida.
Sigui com sigui, no podem pensar en el càncer com una malaltia de causa única, sinó més aviat com el resultat final d’una interacció de múltiples factors, entre els quals s’inclouen l’ambient, els hàbits dietètics, l’herència genètica, etc.
En l’actualitat es realitzen infinitat d’estudis epidemiològics que tracten de buscar associacions de tota índole amb el càncer. Així, per exemple, per discernir entre genètica i ambient, hi ha estudis que comparen la incidència de diferents càncers en una població d’origen amb la incidència d’aquests càncers en una població emigrant en un altre ambient (càncer d’estómac al Japó amb càncer d’estómac en successives poblacions d’emigrants japonesos als Estats Units).

Morfologia del càncer
La diferenciació cel·lular d’un tumor és el grau en què les cèl·lules canceroses s’assemblen a les cèl·lules normals de les que procedeixen, tant morfològica com funcionalment. Les cèl·lules normals que constitueixen l’organisme estan molt diferenciades, la qual cosa els permet realitzar funcions específiques. Generalment, els tumors benignes són ben diferenciats i els càncers varien des de ben diferenciats a indiferenciats.

En general, el que diferencia un càncer maligne d’un altre benigne, és la capacitat que posseeixen les seves cèl·lules d’aconseguir una metastatitzar (la capacitat que té una cèl·lula tumoral d’infiltrar al torrent sanguini o limfàtic). Aquesta característica s’adquireix després de successives alteracions en el material genètic cel·lular ( de 5 a 10 com p.e. pèrdua de gens supressors de tumors com el p53, p21, p16 o el BCl3).
Els gens supressors de tumors són els encarregats de regular el cicle cel·lular i la mort cel·lular programada (apoptosi) respectivament.
L'apoptosi és comparable a un suïcidi, a fi de preservar la integritat cel·lular del teixit conservant només les cèl·lules sanes.
- Augment de la divisió cel·lular que fa que les cèl·lules tumorals es troben en continu divisió.
- Creixement descontrolat: no es deixa influir pels factors de creixement ni altres estímuls externs.
- Metàstasi: (invasió) és la capacitat que tenen les cèl·lules tumorals d’infiltrar o penetrar en els teixits normals, en els vasos sanguinis o limfàtics. Fases:
- Adherència cel·lular: És l’ancoratge de la cèl·lula tumoral mitjançant l’adquisició de receptors específics a la membrana basal i a la matriu extracel·lular
- Mobilitat: És la migració o locomoció de les cèl·lules malignes a través de la matriu cel·lular per a arribar a un vas sanguini o limfàtic i migrar una certa distància per iniciar la formació d’una nova colònia.


Per saber més i entendre millor.
Cicle cel·lular i división cel·lular
Reproducció i meiosi
Herència genètica
Els fills s'assemblen els pares, però no són idèntics. Tots els individus d'una espècie són semblants. Les causes de la semblança i, a la vegada, de la diferència entre organismes en emparentats ens ha preocupat des de sempre. Des de mitjans segle passat s'han anat coneixent els mecanismes pels quals els fills s'assemblen els pares però sensata siguin idèntics, i el coneixement d'aquests mecanismes explicat també la raó per la qual els éssers vius poden anar canviant les seves característiques al llarg del temps.

Està clar que sóm diferents. Sense adonar-nos, primer conexíem el pare i la mare. Després el cercle s'ha anat fent més gran, fins que ara distingim, coneixem a molta gent.
Per distingir aquestes persones observes característiques de la cara de les persones i en reconeixes diferències i semblances.
Aquestes característqiues són trets o caràcters morfològics. Són qualitats biològiques susceptibles de variar.
Així un caràcter de l'espècie humana és el color del seu cabell. I cada persona tendrà una manifestació pròpia d'aquest caràcter, o sigui un color concret (ros, castany obscur, pèl-roig,...)

orelles petitones
mida del peu
color blau dels ulls
l'alçada
pesar 80Kg
No tots els caràcters que presenten els éssers humans són heretables.
Per exemple, no es pot transmetre als fills l'habilitat de jugar a futbol, ni el domini d'un idioma o de tocar el piano,...
Tots aquests caràcters són adquirits al llarg de la vida, i aquests no s'hereten.
La influència de l'ambient.
El fet d'expressar-se en un idioma o en un altre, és una característica que no s'hereta, depèn només de l'ambient.
La forma de les ungles té una base hereditària sobre la qual poden influir diferents característiques ambientals, com per exeemple la feina. És un caràcter determinat genèticament en què l'ambient pot afectar a la seva manifestació. En tot cas, mai es transmet la modificació ambiental.
El grup sanguini és un caràcter estrictament genètic, s'hereta dels pares i no pateix cap modificació ambiental. En aquests darrers anys ha sorgit una branca de la genètica, l'epigenètica, que estudia com l'ambient i la història de l'individu influeixen sobre l'expressió dels gens.
Fenotip = Genotip + Ambient
Aquest personatge asiàtic té totes les característqiues tòpiques: baixet, groc, ulls allargats, pòmuls sortits i amb les dents grosses.
Però segur que tenen aquestes trets? La població del Japó, en el segle passat, vivia amb una dieta basada amb l'arròs. Aquesta dieta té un fort dèficit vitamínic, per això el creixement era escàs i la coloració de la pell groguenca.
En un segle no ha canviat els gens el que ha canviat ha estat l'ambient i ha fet que la manifestació del caràcter també canviï.

Fenotip: cada una de les manifestacions d'un caràcter, és el ressultat de la interacció de tota la nostra informació genètica (gens) amb un ambient determinat.
Genotip i el seus al·les.
Com hem explicat abans un caràcter presenta diferents manifestacions.
Així un gen tendrà diferents forme, ja que cada fenotip s'ha de correspondre amb una forma diferenta del gen.
Cada forma diferents que pot tenir un gen se l'anomena al·lel.
En les espècies diploides cada filament d'ADN està duplicat, una ve del pare i l'altre ve de la mare (filements que donen lloc als cromosomes homòlegs). Per tant cada gen està també dues vegades, un en cada cadena, per la qual cosa cada caràcter està determinat per l'acció de dos al·les, que poden ser igual o diferents.
Quan els dos al·les per un gen són iguals al individu se l'anomena homozigot o raça pura.
Si els dos al·lels són diferents es parla d'individus heterozigots o híbrids.
La posició que ocupa un gen en un cromosoma és el seu locus, i com cada gen es troba en dos cromosomes és el seu loci (plural de locus)
Relacions entre al·lels.
Segons la relació que tenen els al·lel entre ells, tenim els diferents tipus d'herència:
- Dominant i recessiu
- Codominància
- Herència intermèdia o dominància incompleta
- Al·lelisme múltiple
- Herència poligènica
- Lligada al sexe
- Influïda pel sexe
- Epistàsies...
1. Mendel
 Gregor Mendel (Austría, 1822-1884) fou un religiós agustí i naturalista, professor de ciències naturals a l'escola primària de Brno, que va realitzar experiments d'hibridació cultivant pèsols, fent encreuaments per pol·linització artificial. Tot això ho va publicar en la seva obra de 1865 “Versuche über Plflanzenhybriden” (“Experiments d'hibridació en plantes”), on descrigué la transmissió de caràcters d'una generació a una altra en algunes espècies i varietats de Pisum (pesoleres). En aquell moment no tingué pràcticament cap impacte. Malgrat algunes citacions, no fou fins al 1900, setze anys després de la mort de Mendel, que el seu treball fou redescobert de forma simultània per Hugo de Vries, Correns i Tschermak, divulgant la seva obra, des de llavors és considerat pare de la genètica.
Gregor Mendel (Austría, 1822-1884) fou un religiós agustí i naturalista, professor de ciències naturals a l'escola primària de Brno, que va realitzar experiments d'hibridació cultivant pèsols, fent encreuaments per pol·linització artificial. Tot això ho va publicar en la seva obra de 1865 “Versuche über Plflanzenhybriden” (“Experiments d'hibridació en plantes”), on descrigué la transmissió de caràcters d'una generació a una altra en algunes espècies i varietats de Pisum (pesoleres). En aquell moment no tingué pràcticament cap impacte. Malgrat algunes citacions, no fou fins al 1900, setze anys després de la mort de Mendel, que el seu treball fou redescobert de forma simultània per Hugo de Vries, Correns i Tschermak, divulgant la seva obra, des de llavors és considerat pare de la genètica.
Mendel, va utilitzar com a maetrial la pesolera comú (Pisum sativum). La tria d'aqquesta planta per a les investigacions va resultar ideal per diversos motius:
- És una planta bona de conrear i de creixement ràpid, per la qual cosa en poc temps se'n poden estudiar diverses generacions.
- Presenta varietats amb set característqiues fàcilment observables, que es presenten en dues formes possibles.
- Permet controlar-ne la polinització, ja que les flors són hermafrodites. Per això, la pesolera tant es pot autofecundar com se'n pot fer una fecundació creuada de manera artificial.
A més el mètode utilitzat per Mendel (estudiar un caràcter o dos com a màxim en cada experiment, estudiar la descendència després de diverses generacions, utilitzar l'anàlisi matemàtic per demostrar el resultats) li van permetre arribar a resultats coherents amb les seves hipòtesis.

2. Lleis de Mendel
1r llei. Principi d'uniformiat. 2n llei. Principi de segregració.
3r llei. Principi de la segregració independent.
 - Contenidos. Las leyes de la herencia. Reproducción y herencia. Leyes de Mendel.
- Contenidos. Las leyes de la herencia. Reproducción y herencia. Leyes de Mendel.
- Actividades.
Exercicis resolts:
1 - 28 problemas de genética resueltos.
2- Cajón de ciencias. Problemas de genética resultos.
3- Practica con 99 problemas de genética resueltos.
3. Herència intermèdia o Dominància incompleta.
Dos individus homozigots diferents, quan s'uneixen, tenen com a resultat heterozigots amb característqiues intermèdies dels dos progenitors.

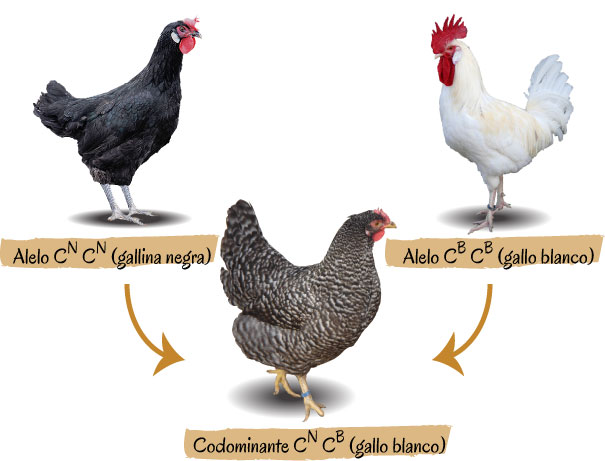
4. Codominància.
Dos individus homozigots diferents, quan s'uneixen, tene com a resultat heterozigots amb les dues característqiues del dos progenitors.
5. Al·lelisme múltiple.

6. Herència poligènica.
Però hi ha moltes altres característiques que són el resultat de l'acció acumulativa de molts de gens i la influència de l'ambient. (alçada, pes, color de la pell,...)
Aquestes gens donen lloc a la gran diversitat d'individus en cada espècie.
7. Herència lligada al sexe.
El cromosoma X és més gran que el cromosoma Y, i té molts més gens.
- En els homes, com que només tenen un cromosoma X, només tenen un al·lel i per tant s'expresserà sempre.
- En les dones, que tenen dos cromosomes X, podran ser: homozigotes dominats, homozigotes recessives i si són heterozigotes s'anomenen portadores.


8. Herència influïda pel sexe.
Aquest tipus d'herència, generalment, es deu a l'acció de les hormones sexuals pròpies de cada sexe.
Així, hi ha al·lels que en els homes es comporten com a dominats i en les dones, com a recessius, o viceverse.
c: gen per a la calvície

9. Teoria cromosòmica de l'herència.
Punts de la teoria cromosòmica de l'herència:
- Els gens es troben als cromosomes, col·locats l'un a continuació de l'altre.
- En un cromosoma hi ha molts de gens. Els gens que es troben molts junts tendeixen a heretar-se junts, s'anomenen gens lligats.
- Els gens d'un mateix cromosoma es poden heretar per seperat a causa de l'entrecreuamnet genètic que té lloc durant la meiosi entre cromàtides homòlegues.
 Multimèdia
Multimèdia
Erase una vez los inventores
La informació i la manipulació genètica
1. L'ADN
1.1 Els àcids nucleics
- desoxirribosa - ADN

- A - Adenina
Pirimidíniques
- T - Timina - ADN

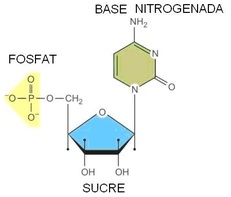

La unió d'un sucre + base nitrogenada + grup fosfat = nucleòtid.
La unió entre nucleòtids es fa mitjançant l'enllaç fosfodiéster.


Va obrir al camí a la comprensió en termes moleculars de l’herència i de la forma d’actuar dels gens.
Però abans de conèixer l’estructura de l’ADN, anem a resumir el que es sabia sobre els gens i sobre l’ADN abans que Watson i Crick comencessin el seu treball:
- Es sabia que els gens – factors hereditaris – descrits per Mendel, estaven associats a caràcters específics, però la seva naturalesa física era desconeguda.
- Al 1871, Miescher, va descobrir que el nucli de les cèl·lules contenien una substància àcida que va anomenar àcid nucleic.
- Es sabia que els gens estaven situats en els cromosomes.
- Es va descobrir que els cromosomes estaven formats per ADN i proteïnes.
- Al 1928, Frederick Griffith i més tard al 1944, Avery, McLeod i McCarty , van demostrar que l’ADN és la partícula portadora de la informació genètica.
- La teoria “un gen – un enzim” defensava que els gens controlaven l’estructura de les proteïnes.
1.2. Experiment de Griffith
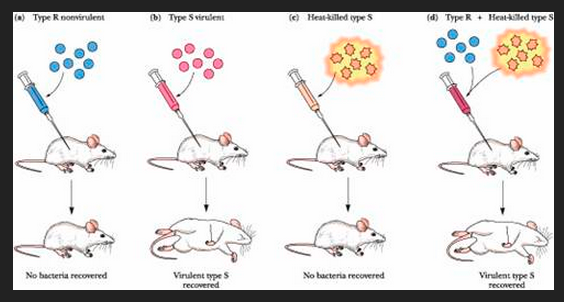 L'experiment de Griffith (1928), fou un dels primers experiments que suggeriren que els bacteris eren capaços de transferir informació genètica a través del procés de transformació bacteriana.
L'experiment de Griffith (1928), fou un dels primers experiments que suggeriren que els bacteris eren capaços de transferir informació genètica a través del procés de transformació bacteriana.
Griffith usà dues soques de pneumococ amb les que infectà ratolins, una soca tipus-S (smooth, llís) i l'altra un tipus-R (rough rugós).
La soca -S es recobreix ella mateixa amb una càpsula bacteriana que la protegeix del sistema immunitari de l'hoste, resultant en la mort de l'hoste, mentre que la soca -R no disposa d'aquesta càpsula protectora i és rebutjada pel sistema immunitari de l'hoste, i, per tant, és menys virulenta.
En l'experiment els bacteris de la soca virulenta -S foren morts per calor i les restes foren afegides en un cultiu de la soca -R. En afegir les restes de bacteris -S morts a bacteris -R vius, aquests darrers aconseguien la capacitat de matar el ratolí en ésser-li inoculats. Griffith conclogué que algun "principi de transformació" s'havia després de les restes de bacteris de la soca virulenta -S morta i havia convertit els bacteris -R en bacteris virulents -S.
Avui en dia, es coneix que el "principi transformant" de Griffith és l'ADN.
La natura exacta del principi transformant fou verificada pels experiments d'Avery, McLeod i McCarty.
1.3. Experiment de Avery
 L'experiment d'Avery i dels seus col·laboradors Colin MacLeod i Maclyn McCarty es va realitzar l'any 1943. Mitjançant aquest experiment els científics reixiren a demostrar que el considerat principi trasformant (o sigui el portador de la informació gènica) descobert el 1928 per Griffith era l'ADN.
L'experiment d'Avery i dels seus col·laboradors Colin MacLeod i Maclyn McCarty es va realitzar l'any 1943. Mitjançant aquest experiment els científics reixiren a demostrar que el considerat principi trasformant (o sigui el portador de la informació gènica) descobert el 1928 per Griffith era l'ADN.
 Griffith proposà que dins l'interior d'una mescla que contingui bacteris morts S i bacteris vius R, ha d'haver hagut un bescanvi d'alguna substància (el material genètic) que hauria conferit virulència als bacteris R (que s'haurien transformat en els S).
Griffith proposà que dins l'interior d'una mescla que contingui bacteris morts S i bacteris vius R, ha d'haver hagut un bescanvi d'alguna substància (el material genètic) que hauria conferit virulència als bacteris R (que s'haurien transformat en els S).
L'experiment d'Avery va tractar de determinar quina era aquesta substància.
Avery va fer un cultiu dels pneumococcus del tipus S. En va trencar les cèl·lules (trencant la membrana cel·lular) per obtenir una solució o extracte cel·lular.
Van aconseguir separar l'extracte cel·lular dels seus components macromoleculars i seguidament van intentar entendre quines d'aquestes substàncies podien transformar els bacteris R avirulents en bacteris S virulents. Els animals de laboratori (ratolins) van sobreviure quan van ser tractats amb totes les biomolècules a les quals s'havien retirat els àcids nucleics: per tant el material genètic havia de ser ADN o ARN.
Per esbrinar quina de les dues substàncies era es van tractar les mostres amb dos enzims diferents un d'ells degradava l'ARN i l'altra degradava l'ADN i així es va determinar que el material genètic havia de ser l'ADN.
1.4. La teoria un gen - un enzim
 La teoria més ampliament acceptada sobre la manera d'actuar dels proveen dels treballs dels genetistes Beadle i Tatum, amb le moho vermell del pa Neurosporo crassa.
La teoria més ampliament acceptada sobre la manera d'actuar dels proveen dels treballs dels genetistes Beadle i Tatum, amb le moho vermell del pa Neurosporo crassa.  Varen sotmetre la floridura a diferents radiacions per originar individus mutants incapaços de sintetitzar determinats compostos. Aquests organismes defectuosos només sobrevivien si s'afegia al medi el compost corresponent. Van demostrar que els individus mutants tenien un gen afectat, i que això impedia la formació d'un enzim.
Varen sotmetre la floridura a diferents radiacions per originar individus mutants incapaços de sintetitzar determinats compostos. Aquests organismes defectuosos només sobrevivien si s'afegia al medi el compost corresponent. Van demostrar que els individus mutants tenien un gen afectat, i que això impedia la formació d'un enzim. Un gen conté la informació perquè els aminoàcids s'uneixin en un determinat ordre i formin un enzim.
1.5. La doble hèlix
Després de demostrar el paper tan important de l’ADN en l’herència, molts científics es van disposar a determinar la seva estructura amb exactitud.
Per què... Com podia ser que una molècula amb tants pocs elements diferents pogués emmagatzemar la immensa varietat informativa de les estructures de totes les proteïnes dels éssers vius?
Els primers que van tenir èxit en trobar una estructura raonable per l’ADN van ser:
James Watson i Francis Crick al 1953

Per dur a terme el seu treball tenien algunes pistes:
- La Þ-hèlix de proteïnes obtinguda per Linus Pauling.
- L’estructura de l’ADN obtinguda per difracció de Raigs X de Rosalind Franklin
- El treball realitzat per Erwing Chargaff
- Linus Pauling
 Va ser un dels primers científics a aplicar els principis de la mecànica quàntica per a donar explicació als fenòmens de difracció dels raigs X i va assolir descriure satisfactòriament les distàncies i els angles dels enllaços químics entre àtoms de diverses molècules.
Va ser un dels primers científics a aplicar els principis de la mecànica quàntica per a donar explicació als fenòmens de difracció dels raigs X i va assolir descriure satisfactòriament les distàncies i els angles dels enllaços químics entre àtoms de diverses molècules.El seu treball al costat del químic nord-americà Robert B. Corey el va dur a reconèixer l'estructura helicoïdal de certes proteïnes i va realitzar intents fallits de descriure l'ADN.
- Rosalind Franklin

 Imatge 51
Imatge 51Rosalind Franklin, una dona de forta personalitat, va mantenir una relació complexa amb en Maurice Wilkins, qui va mostrar sense el seu consentiment les seves imatges de difracció de raigs X de l'ADN a James Watson i a Francis Crick. S'admet que cap altra inspiració fou tan forta com aquesta per la seva publicació, el 1953, de l'estructura de l'ADN.
Aquest descobriment no fou casual, sinó que Franklin va demostrar la seva habilitat per obtenir les millors imatges i per interpretar-les correctament en la investigació d'altres objectes, com l'estructura del grafit o la del virus del mosaic del tabac.
Franklin va morir prematurament, de càncer d'ovari, el 1958 a Londres, amb tota probabilitat per efecte de les repetides exposicions a les radiacions en el curs de les seves investigacions.
- Lleis de Chargaff
 Chargaff, analitzant les bases nitrogenades de l'ADN en diferents formes de vida, va descobrir dues regles que van ajudar la descoberta de la doble hèlix de l'ADN. Lleis de Chargaff:
Chargaff, analitzant les bases nitrogenades de l'ADN en diferents formes de vida, va descobrir dues regles que van ajudar la descoberta de la doble hèlix de l'ADN. Lleis de Chargaff:La quantitat de bases Adenina (A) és igual a la quantitat de Timina (T), i la quantitat de bases Guanina (G) és igual a la quantitat de Citosina (C). De manera que la suma de bases nitrogenades púriques (A i G) és igual a la suma de les pirimidíniques (T i C). Així s'estableix la complementarietat de les bases nitrogenades en l'ADN.
- Segona llei:
La proporció de bases nitrogenades és semblant en els diferents òrgans d'un mateix organisme, però que aquestes proporcions són específiques i diferents per a cada espècie.
Aquests resultats refutaven creences prèvies que afirmaven que l'ADN era la repetició de les quatre bases amb una monotonia i sense cap altra rellevància. Des d'aleshores, la comunitat científica va començar a creure que era en l'ADN i no en les proteïnes on es determinava l'especificitat molecular biològica dels organismes.
ADN
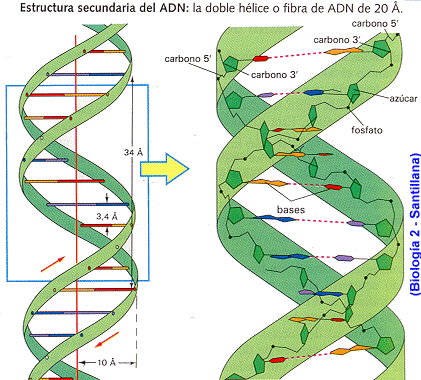
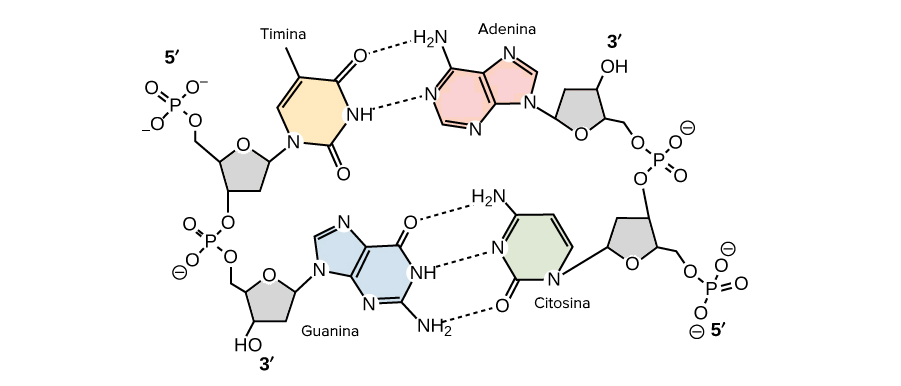
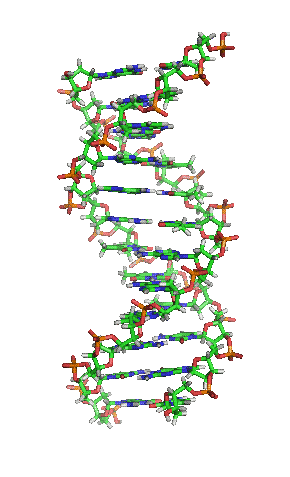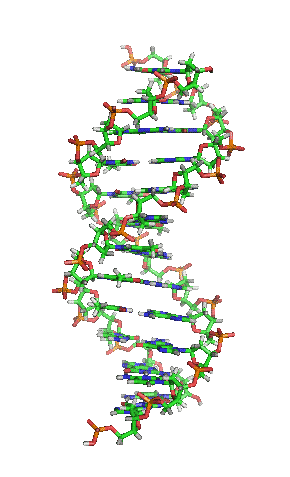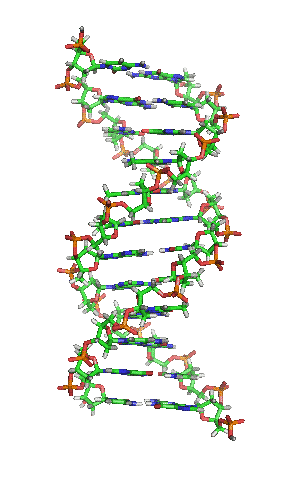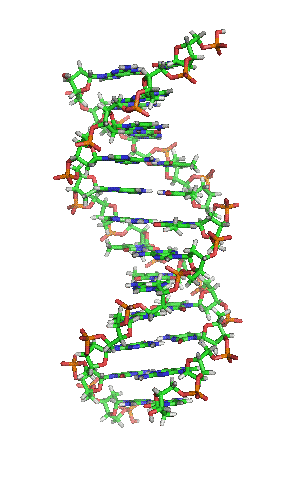 - La molècula d'ADN consisteix en una hèlix doble, ja que està formada per dues cadenes de nucleòtids enrotlladas al llarg d'un eix comú.
- La molècula d'ADN consisteix en una hèlix doble, ja que està formada per dues cadenes de nucleòtids enrotlladas al llarg d'un eix comú. - Aquesta doble hèlix és dextrogira.
- Dóna un gir complet cada 3,4 nm.
- Les dues cadenes es disposen en sentits oposats, és a dir, una en sentit 3' → 5' i l'altra en sentit 5' → 3'. Les dues cadenes són, per tant, antiparal·leles.
- Les bases nitrogenades són a l'interior de la doble hèlix, amb els plans dels seus anells perpendiculars a l'eix de l'hèlix, mentre que els esquelets pentosa-fosfat de les dues cadenes es troben a la part exterior.
- La distància entre cada parell de bases és de 0,34 nm i com que la doble hèlix fa una volta completa cada 3,4 nm, conté 10 parells de bases per volta.
- Les dues cadenes queden unides per ponts d'hidrogen formats entre la base nitrogenada d'una cadena i la de l'altra cadena amb la qual queda enfrontada. Els aparellaments sempre són entre A-T i G-C, és a dir, sempre s'enfronta una base púrica amb una pirimidínica, i mai dues del mateix tipus. El parell A-T pot formar 2 ponts d'hidrogen i el parell G-C en pot formar 3.
Aquest model mostrava ja la possibilitat que l'ADN pogués complir les següents funcions:
- Contenir la informació genètica codificada a la seqüència de bases. Malgrat que l'ADN estigui forma només per quatre tipus de bases, el número de seqüències possibles i, per tant, la diversitat de la informació emmagatzemada, és enorme.
- Reproduir-se exactament per a originar dues molècules filles idèntiques a la molècula original. Això és absolutament necessari per tal que la informació genètica pugui ser transmesa fidelment i es pot aconseguir gràcies al fet que l'aparellament entre les bases sempre és A-T i G-C. D'aquesta manera, si les dues cadenes se separen, cadascuna pot servir de motlle per a la síntesi d'una nova cadena complementària idèntica a aquella de la qual s'ha separat.
- Un canvi en la seqüència (mutació) pot modificar la informació i aquest canvi pot ser transmès a la descendència.
Les dues cadenes que formen la doble hèlix poden ser separades per diferents mètodes, entre ells la calor. El procés de separació de les dues cadenes s'anomena desnaturalització de l'ADN.
Molècula tridimensional de l'ADN
ARN
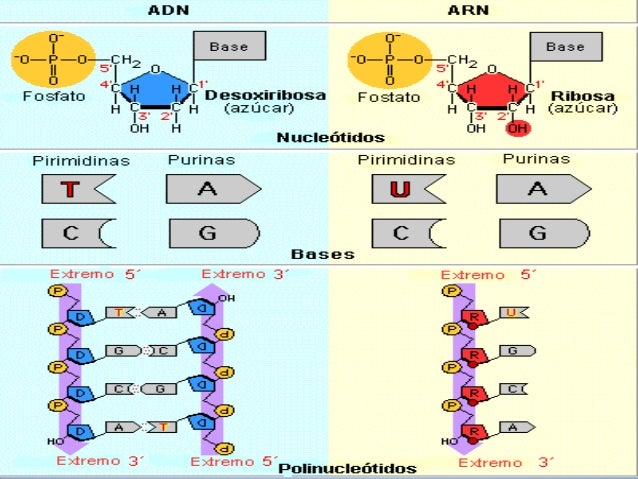 L'ARN (àcid ribonucleic) és un polímer format per la unió de ribonucleòtids d'A, G, C i U mitjançant enllaços fosfodièster.
L'ARN (àcid ribonucleic) és un polímer format per la unió de ribonucleòtids d'A, G, C i U mitjançant enllaços fosfodièster. L'ARN es troba a totes les cèl·lules eucariotes i procariotes així com als virus d'ARN (els virus mai tenen ARN i ADN alhora), tant mono- com bicatenari.
La major part dels ARN són monocatenaris, és a dir, a diferència de l'ADN la molècula està formada per una sola cadena lineal de ribonucleòtids. Això no obstant, poden tenir zones amb estructura de doble hèlix per aparellament de bases complementàries de la mateixa cadena. Els aparellaments en aquest cas són A-U i G-C.
Tan sols un tipus de virus, els retrovirus, posseeixen ARN bicatenari, amb dues cadenes aparellades i estructurades en una doble hèlix.

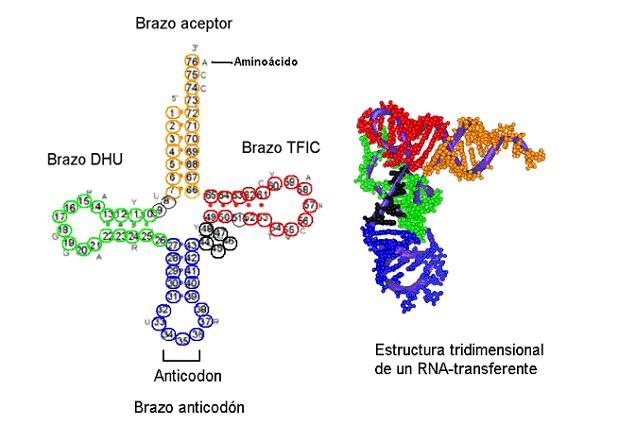 - ARN de transferència (ARNt). Són molècules que contenen entre 70 i 90 nucleòtids.
- ARN de transferència (ARNt). Són molècules que contenen entre 70 i 90 nucleòtids. Estan formats por una única cadena que presenta estructura secundària gràcies a l'aparellament entre bases complementàries en diferents zones de la molècula.
Aquestes regions de doble hèlix deixen entre elles una sèrie de bucles.
Les molècules presenten una conformació tridimensional en forma de L.
La seva funció és captar aminoàcids i transportar-los als ribosomes, tot col·locant-los en el lloc indicat per la seqüència de l'ARNm.
Les quatre zones de doble hèlix delimiten tres bucles un dels quals conté un triplet específic de bases, l'anticodó, que és complementari d'un triplet de bases de l'ARNm al qual s'unirà, el codó. Un ARNt amb un anticodó específic s'uneix sempre al mateix aminoàcid i no a un altre, la qual cosa resulta essencial per a poder traduir la informació transportada des de l'ADN per l'ARNm.
- ARN ribosòmic (ARNr). És el més abundant a les cèl·lules. Es troba formant part dels ribosomes i, com l'ARNt, presenta zones amb doble hèlix.

2. Replicació
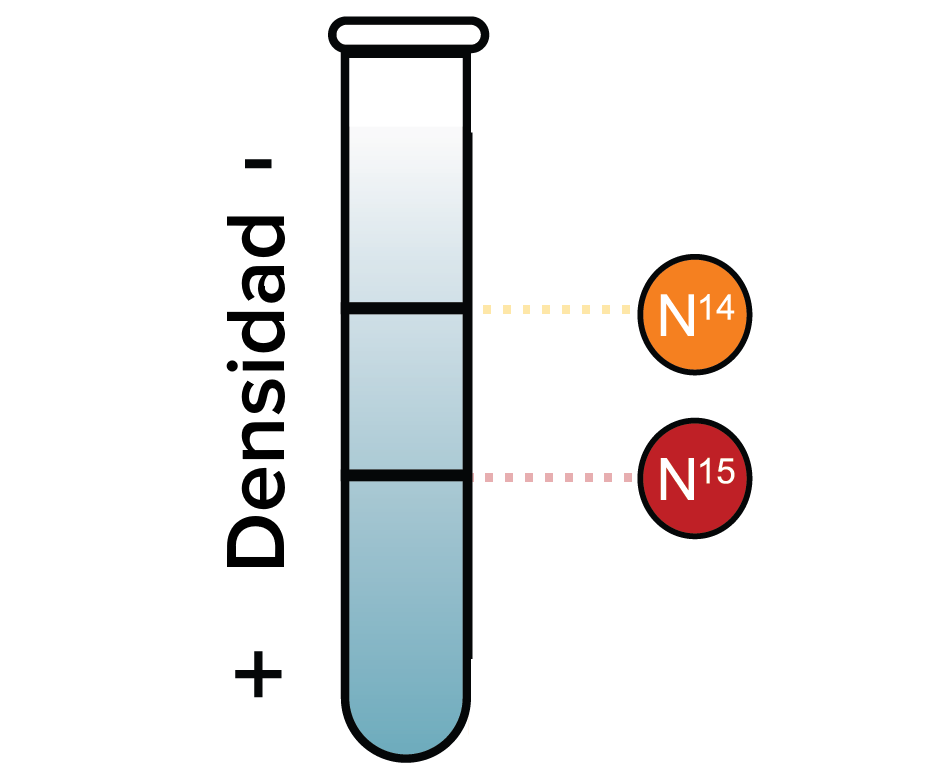
Meselson i Stahl demostraren al 1958 que la replicació de l'ADN és semiconservativa, és a dir, que les dues molècules d'ADN que passen a la generació següent tenen cadascuna una cadena vella i una de nova. Ara bé, com es copia l'ADN per a formar dues noves cadenes idèntiques a la cadena original?




A la replicació de l'ADN intervenen:
-Les cadenes d'ADN en formació van creixent sempre en sentit 5' → 3' mentre que la cadena motlle es copia sempre en sentit 3' → 5'.
- L'ADN polimerasa necessita un extrem 3'-OH lliure, és a dir, un petit fragment de cadena nova per a començar a copiar. Com que aquest enzim no pot sintetitzar-lo, ha d'intervenir un altre enzim diferent que sintetitzi aquest fragment inicial anomenat iniciador o encebador o primer.
- Posseeixen activitat exonucleasa. Quan l'aparellament entre la base de la cadena motlle i la base del nou nucleòtid incorporat no és correcte, el nou extrem no queda correctament situat i l'ADN polimerasa elimina el nucleòtid mal aparellat. Aquesta propietat permet que es corregeixin errades que es podrien produir en la còpia de la nova cadena.
- ARN polimerasa: enzim que sintetitza els iniciadors necessaris, petits fragments d'ARN que ofereixen un extrem lliure per tal que pugui actuar l'ADN polimerasa.
- Topoisomerases i helicases, enzims que possibiliten que la doble hèlix s'obri i que eviten el superenrotllament que es podria produir en anar-se separant les dues cadenes.
- Proteïnes SSB, estabilitzadores de les cadenes senzilles d'ADN.
- ADN lligasa, enzim que uneix extrems 3'-OH i fosfat adjacents dins d'una mateixa cadena, tot consumint ATP.

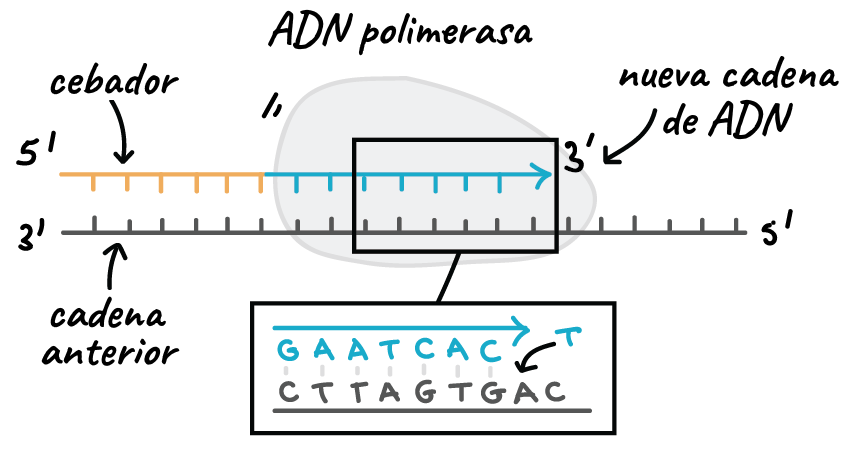

Les dues cadenes d'ADN són antiparal·leles però totes les ADN polimerases conegudes afegeixen nucleòtids en sentit 5' → 3'. D'aquesta manera podria explicar-se la síntesi contínua d'una cadena però no de l'altra.
Al 1968 Reiji Okazaki i Tsuneko Okazaki van demostrar que la cadena que hauria de sintetitzar-se en sentit 3' → 5' se sintetitza en realitat en forma de petits fragments en sentit 5' → 3' d'una manera discontínua. Aquests fragments s'anomenen, en el seu honor, fragments d'Okazaki i quan són complets, els seus extrems són units per l'ADN lligasa.
 D'aquesta manera, l'esquema general de la replicació de l'ADN seria el següent:
D'aquesta manera, l'esquema general de la replicació de l'ADN seria el següent:- Una ARN polimerasa sintetitza en sentit 5' → 3' una petita molècula d'ARN, l'iniciador. A la cadena de síntesi contínua només és necessari un iniciador; a l'altra cadena se sintetitza un iniciador cada 1.000 - 2.000 nucleòtids.
- L'ADN polimerasa va allargant la cadena incorporant-hi desoxiribonucleòtids a l'extrem 3'-OH lliure. Els nucleòtids s'hi incorporen seguint les regles d'aparellament A-T i G-C. Si un aparellament no és correcte, l'ADN polimerasa elimina el nucleòtid incorrecte. A la cadena de síntesi discontínua, quan l'ADN polimerasa arriba al fragment d'ARN iniciador anterior, l'elimina i omple el forat amb desoxiribonucleòtids.
- Finalment l'ADN lligasa uneix els extrems dels fragments d'Okazaki.
De fet, a partir d'un origen de replicació la síntesi és bidireccional en tots els organismes, de manera que tenim dues bombollrd de replicació que van avançant en sentits oposats. Als procariotes hi ha un sol origen de replicació, mentre que als organismes eucariotes hi ha molts, i les diverses forquilles de replicació van avançant fins que es troben amb una altra forquilla de replicació o amb l'extrem de la molècula.
Vols veure el procés en marxa?
3. Expressió gènica


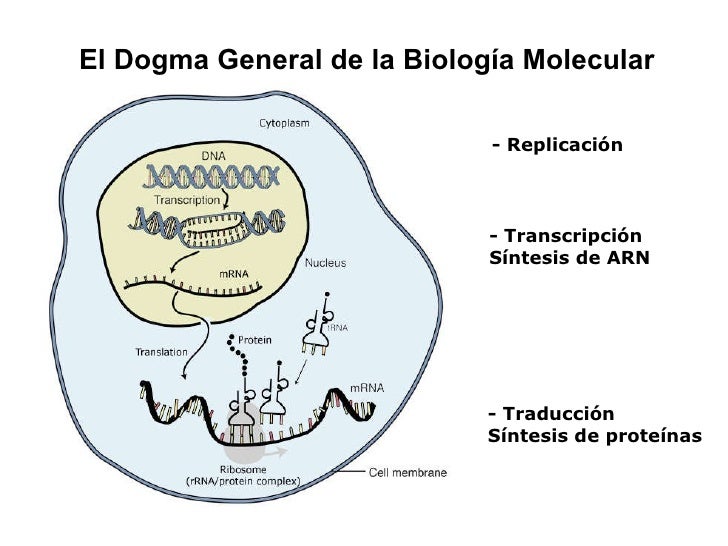
Dogma central de la biologia molecular
La informació genètica (ADN) és un missatge xifrat.
El missatge s'ha de descodificar (ARN) - Transcripció.
Per a convertir-se finalment en una proteïna - Traducció
3.1. Transcripció
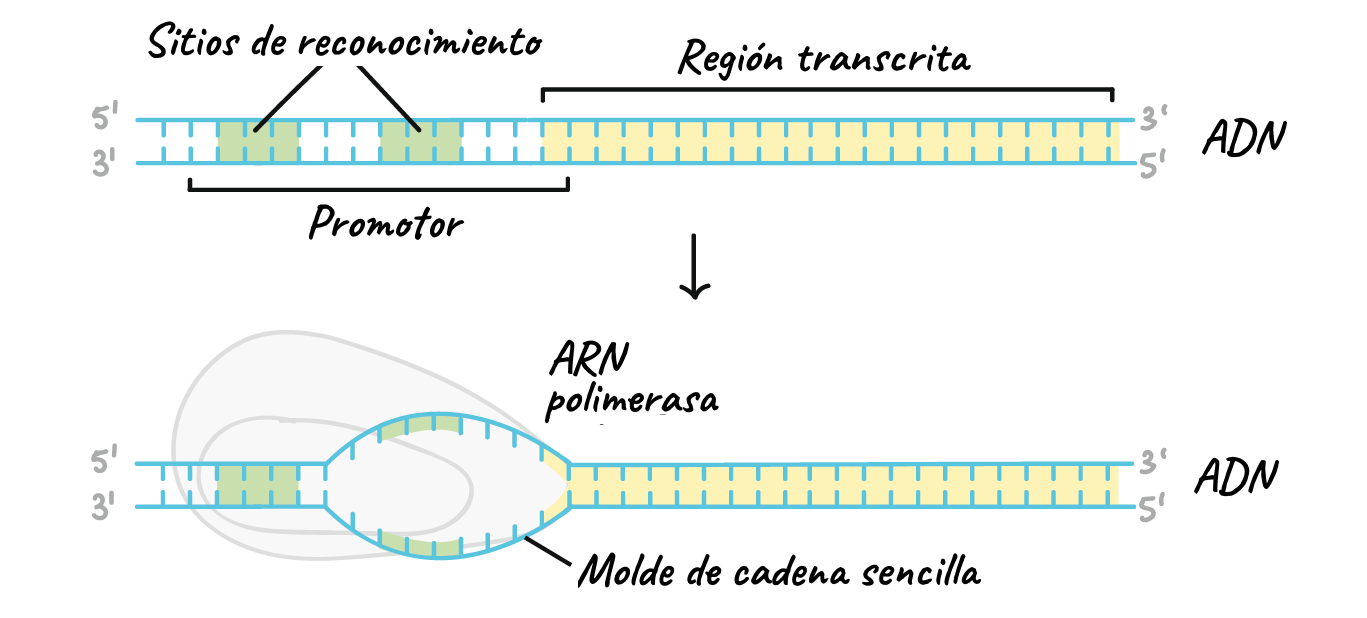
Durant la transcripció genètica, les seqüències d'ADN són copiades a ARN mitjançant un enzim anomenat ARN polimerasa.
Aquest procés té lloc en el nucli de la cèl·lula.

- Reconeix un lloc d'inici per la transcripció, l'inici del gen.
- Desenrotlla parcialment la molècula d'ADN, activitat helicasa.
- Sintetitza un l'ARNm seguit les regles de complementarietat de bases, amb l'excepció que la Timina és substituïda per Uracil (G-C /A-U).
- Acaba la cadena i enrrotlla l'ADN.
 2.
2.
3.

4.

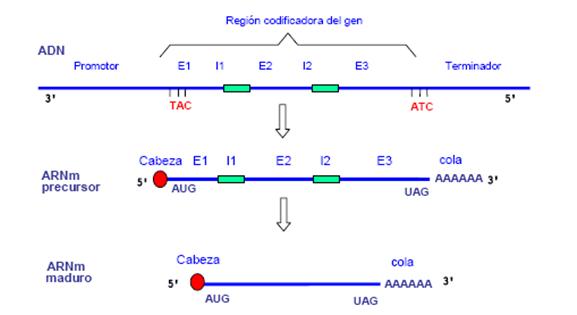
1. Gen: és una seqüència lineal de nucleòtids d'ADN. És la unitat d'emmagatzemament d'informació i unitat d'herència en transmetre aquesta informació a la descendència.
2. Triplet: seqüència de tres nucleòtids en un àcid nucleic i en particular en un ARN missatger.
Codó: Els triplets de l'ADN reben el nom de codó.
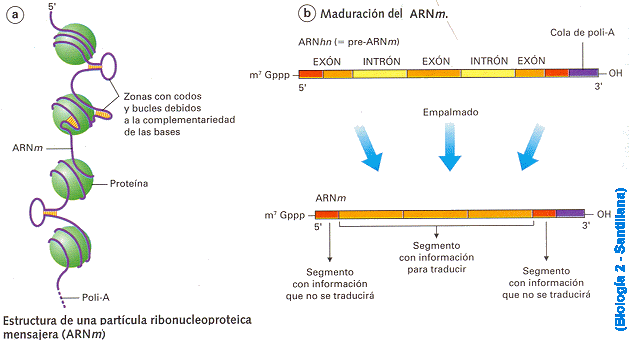
3: Exó: és cada un dels segments d'ADN codificants d'un gen que contenen informació i no són eliminats en la maduració de l'ARNm.
Intró: segments d'ADN que no contenen informació i sí que són eliminats durant la maduració.
4. Maduració - Splicing: Eliminació dels introns, addició de un CAP (7-metil guanosina) i una coa de poli-A.
3.2. Traducció

En els eucariotes la traducció ocorren en el citoplasma o en la membrana del RER.
Així:
- L'ARNm fabricat surt del nucli a través dels porus nuclears i s'ajunta als ribosomes.
- Els ribosomes llegeixen el misstge en grups de tres nucleòtids - codó.
- L'ARNt porta els aminoàcids, un de diferent per a cada anticodó.
- L'ARNt col·loca el seu aa als codons del ARNm per complementarietat de bases amb el seu anticodó.
- Els aa es van enllaçant a mesura que el ribosoma avança.
- El ribosoma troba un codó de STOP i es desfà. La proteïna està formada.


 2.
2.
3.3. Codi genètic
 L'ADN havia d'especificar, d'alguna manera, la seqüència d'aminoàcids de les proteïnes.
L'ADN havia d'especificar, d'alguna manera, la seqüència d'aminoàcids de les proteïnes.
Necessàriament havia d'haver algun codi que traduís el missatge de l'ADN en forma de seqüència d'aminoàcids a la proteïna.
Nirenberg al 1961.
Característiques del codi Genètic:
- La informació es llegeix en grups de tres nucleòtids, anomenats triplets. El codi havia de tenir tres lletres com a mínim (si fos de dues lletres -o nucleòtids- només hi hauria 4^2=16 possibles combinacions, insuficients per a codificar els 20 aminoàcids que formen les proteïnes; amb tres nucleòtids hi ha 4^3=64 combinacions). - Com que hi ha 64 triplets i 20 aminoàcids és possible que hi hagi aminoàcids codificats per més d'un triplet. Per això diem que el codi és degenerat.
- El codi és seqüencial i no encavalcat: els nucleòtids es llegeixen un rere l'altre en grups de tres i cada triplet segueix l'anterior sense encavalcar-se amb ell.
- El codi té triplets sense sentit que no codifiquen aminoàcids sinó senyals d'aturada.
- El codi no és ambigu: un triplet codifica un sol aminoàcid i mai més d'un (tot i que un aminoàcid pugui ésser codificat per més d'un triplet).
- El codi és universal, vàlid per a tots els éssers vius. Posteriorment es va trobar alguna excepció en el cas de l'ADN mitocondrial, on el triplet UAG codifica Trp (triptofà) en comptes de ser un triplet d'aturada o AUA codifica Met (metionina) en comptes d'Ile (isoleucina).
 |
|
|
Per saber més i entendre millor
Van expossar a l'acció dels raigs UV algunes espores sexuals que provenen de cert tipus d'aparellament de Neurospora. Després van deixar germinar l'espores en un medi complet, és dir, enriquit amb les vitamines i aminoàcid. Una vegada desenvolupat el miceli, es van fer creuaments amb altres tipus d'aparellement. Una vegada que van crèixer, es van col·locar porcions de miceli de cada cultiu en un medi mínim. De vegades el creixement continuava, a vegades es suspenia; quan això passava aquella raça particular rebia vitamines, aminoàcids, entre altres. Fins aconseguir el creixement. Finalment es pot establir que cada raça deficient era capaç de crèixer en un medi mínim, al qual 'havia afegit una substància accesoria, per exemple la Timina.
Beadle i Tatum van suposar que la radiació UV havia produït una mutació del gen que possibilita la síntesis de la tiamina, i havia transformat en un al·lel que no era capaç de fer-ho.
La síntesi de tiamina a partir de les susbtàncies simples presents en el medi mínim no ocorre mitjançant una sola reacció química, sino amb una sèrie completa de reaccions. Com totes les reaccions químiques en el éssers vius, cada una requereix la presència d'un enzim específic mitjançant l'addicció de compostos intermedis (precursors) al medi en el qual creixia el moho. Aquest experiment va permetre localitzar exactament quina fase en la síntesi de la tiamina està bloquejada en la raça mutant.
Amb aquesta base, van crear la teoria d'un gen - un enzim referent a l'acció del gen, que pot formular-se en els següents termes: cada gen en un determinat organisme regula la producció d'un enzim específic.
A començament dels anys 50 es va veure que la insulina estava formada per una cadena lineal d'aminoàcids que tenien un ordre definit. Si la insulina tenia una seqüència definida, genèticament determinada, el més probable és que la resta de proteïnes també tinguessin una seqüència fixa, diferent per a cada proteïna.
Al 1961 per Nirenberg. Van posar ARN poliuracil (una cadena d'uracils, UUUUUUU...) a 20 tubs d'assaig. A cada tub d'assaig hi van afegir extracte citoplasmàtic d'Escherichia coli i un aminoàcid marcat radioactivament, diferent a cada un dels tubs. Van deixar un temps perquè hi tingués lloc la reacció i tot seguit van separar els ribosomes de cada tub, a veure què havia passat. Va resultar que els ribosomes del tub que contenia Phe (fenilalanina) estaven marcats radioactivament, és a dir, tenien Phe unida. Per tant, el conjunt de tres nucleòtids UUU codificava l'aminoàcid Phe.
Dogma central
Multimèdia
![]() James Watson explains DNA basepairing
James Watson explains DNA basepairing
![]() Watson, Crick i Franklin
Watson, Crick i Franklin
![]() Duplicación del ADN
Duplicación del ADN
![]() How DNA copies itself
How DNA copies itself
![]() Síntesis de proteinas (Transcripción y Traducción)
Síntesis de proteinas (Transcripción y Traducción)
![]() From DNA to Protein
From DNA to Protein
![]() Somos lo que comemos - Epigenética y genes
Somos lo que comemos - Epigenética y genes
Mutacions
Aquestes alteracions o mutacions provoquen un canvi en l´estructura o en la composició del material gènic.
Aquestes alteracions són accidentals i a l´atzar.
Normalment les mutacions suposen deficiències i poden arribar a ser letals (per la cèl·lula). Però en general són recessives i queden amagades sense causar cap efecte dolent en l´individu, i suposa un aspecte positiu per l´espècie ja que aporta variabilitat. Així un canvi en l´ambient, advers pels individus normals o sense una mutació específica, pot fer que l´individu mutant sobrevisqui a l´ambient.
Exemples:
- Una mutació deficient en un gen específic que controla la replicació (p53) farà que la cel es divideixi sense aturar-se i si no hi ha cap remei (procés de reparació efectiu o de mort cel) pot aparèixer càncer.
- Una mutació recessiva: hemocromatosi hereditària. Malaltia per una mutació puntual en la prot HFE (prot de 343 aa, cr 6: G -> A: Cys -> Tyr ).
Provoca un transtorn del metabolisme del ferro: hi ha una absorció excessiva de Fe que es deposita en diferents òrgans com el fetge i provoca: càncer, cirrosis, esterilitat, pigmentació excessiva de la pell...
La malaltia és asintomàtica en els primers anys de vida ja que les necessitats de Fe són altes i els símptomes començen aparèixer als 45 anys. Es sap que aquesta mutació és originària del nord d´Europa perquè és on es dóna en major freqüencia, i es va distribuir amb les migracions celtes. A Mallorca 1/200 la pateix.
Es creu que aquesta freqüencia tan alta és per què en generacions anteriors a les nostres era freqüent la deficiència de Fe en la dirta per tant ser heterocigot (tenir aquesta mutació) era beneficiós.
Hem dit que les mutacions solen ser recessives i que queden amagades ja que queden amb heterocigosi i com tenim una altra còpia de ADN que està intacta no hi ha cap deficiència visible. Aquestes mutacions es van heredant i passen de generació a generació. Per tant els membres d’una mateixa família tenen mutacions en els mateixos gens. Aquesta és la raó de perquè no es recomanable que els parents tenguin fills, ja que una mutació que en els dos indivudus està en heterocigosis en la seva descedència pot passar a ser homocigot en aquella mutació i provocar una determinada malaltia.
Tipus de mutacions:
Segons el seu origen:
- Espontànies: degudes a la mateixa naturalesa química del ADN
- Induïdes o provocades pels agents mutàgens
Segons les cèl·lules afectades:
1.Per un error en els mecanismes moleculars de replicació. (La replicació té sistemes de correcció d´aquests errors)
- Mutació cromosòmica: per un error en l´estructura del cromosoma.
Agents mutàgens
Un mutàgen seria qualsevol substància o radiació que interacciona o reacciona amb el ADN i el pot fer canviar.
Hi ha mutàgens:
- químics: molècules que modifiquen les bases nitrogenades
- químics: molècules que són anàlogues a les bases nitrogenades i les substitueixen
- químics: molècules de intercalació
- físics: radiacions no ionitzants
- físics: radiacions ionitzants
-
Químics
1- Molècules que modifiquen les bases nitrogenades
- Àcid nitròs: la A (adenina) es transforma en hipoxantina que s´aparea amb la C (citosina), això pot provocar un canvi en la pauta de lectura.
 Les nitrosaminas són substàncies que les trobam en el fum del tabac o en els aliments cremats.
Les nitrosaminas són substàncies que les trobam en el fum del tabac o en els aliments cremats.
També els nitrits, que trobam en embutits o aigües contaminades per adobs, poden donar lloc, una vegada metabolitzats, a nitrosamines.
Les nitrosamines provoquen metilacions (-CH3) en les bases nitrogenades. Si es metila el ARN no es molt greu ja que tan sols s’altera l’expressió d’una proteína. Però si es metila el ADN que és la font mare d´informació pot provocar problemes més greus.
Normalment la BN més susceptible de ser metilada és la G que en la pauta de lectura és substituida per una T. La hipermetilació provoca la no expresió d´un gen.
- Agents alquilants: com el gas mostasa, utilitzat en la guerra del Vietnam. Afageix grups acil i provoca conseqüencies semblants a les nitrosamines.

I altres com el benzopiré ( que el trobam en el fum del tabac o en els aliments cremats), flavonids ( presents en el cafè, te, cacau, cervesa), derivats piròlics – derivats del tritòfen (el trobam en olis sotmesos a fritures succesives o en aliments sotmesos a temperatures superiors a 200ºC) excés de glucosa ( provoca la glucosilació de les BN del ADN que dificulta la lectura).
2- Molècules que són semblants a les bases nitrogenades i les substitueixen.
El 5-bromouracil pot substituir a una Timina.
El 2-aminopurina pot substituir a una Adenina.
Les modificacions implica aparellaments amb bases diferentes a les complementaries.


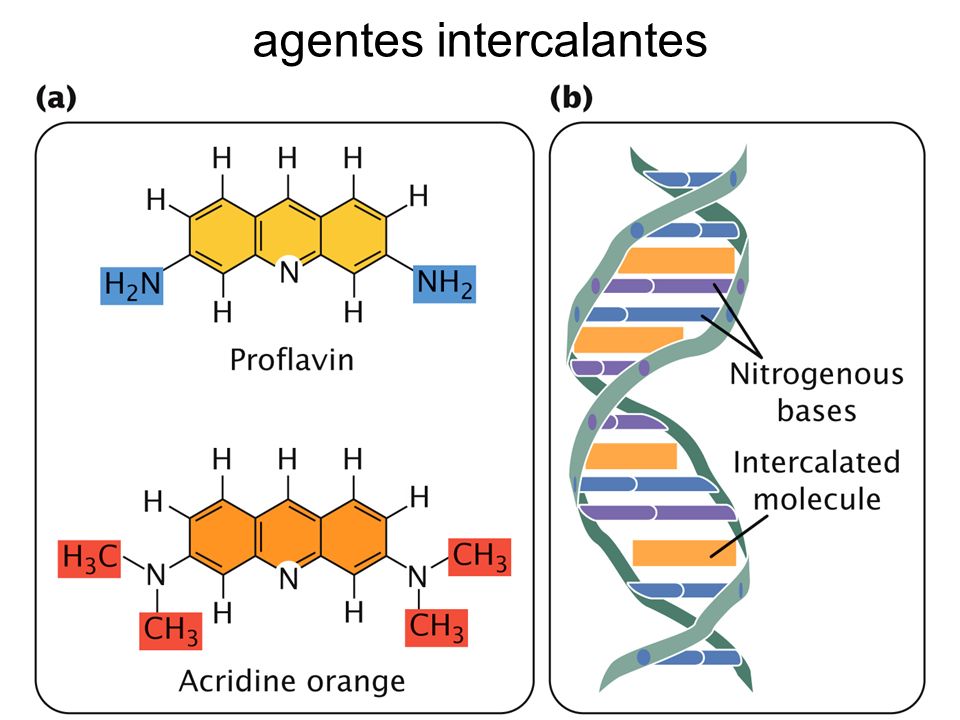
3- Molècules d'intercalació.
Molècules similars a un parell de bases nitrogenades enllaçades, com la acridina o proflavina, que s´introdueixen entre els parells de BN.
Quan es produeix la duplicació o replicació, poden aparèixer insercions o delecions d’un parell de bases nitrogenades i així la pauta de lectura es canvia.
-
Físics
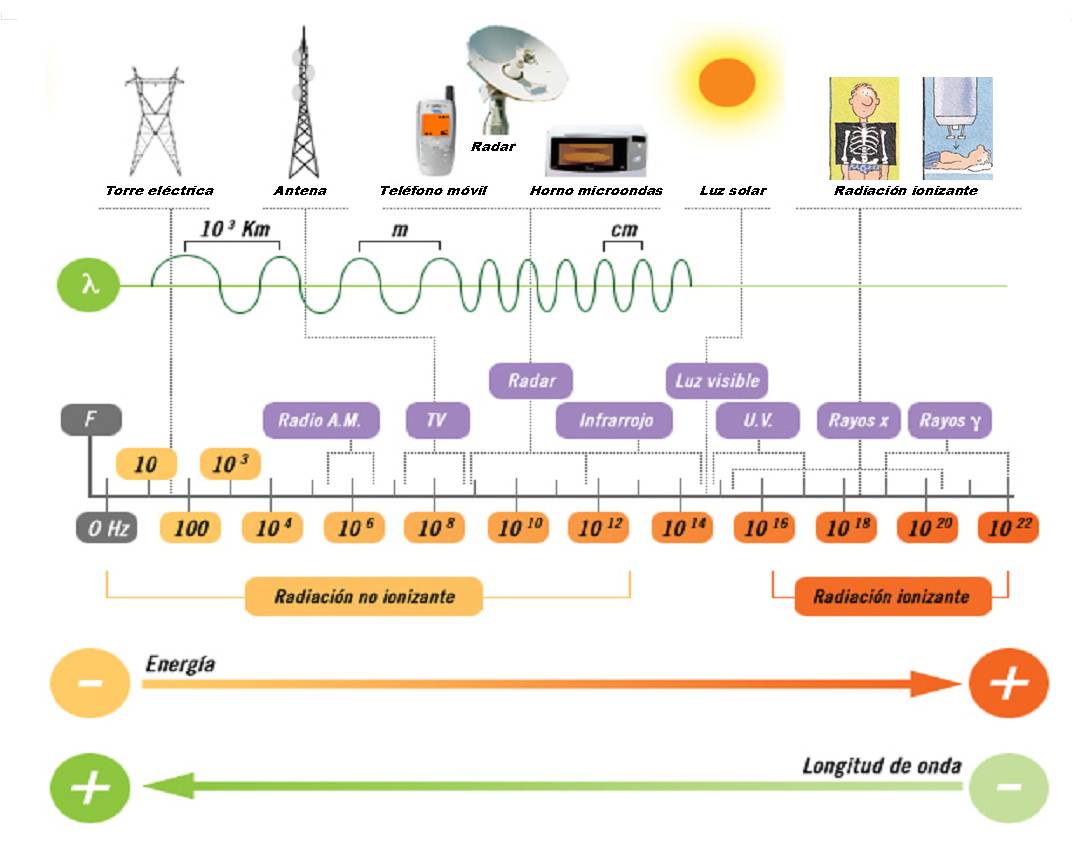
1. Radiacions no ionitzants
Són radiacions electromagnétiques, com la llum, però de menor longitud d´ona (entre 160 – 400 nm: UV) i per tant més energétiques.
Aquestes radiacions són absorbides pel ADN i provoca el bot d’alguns electrons a nivells energètics superiors, afavorint la formació d’enllaços covalents, entre dues pirimidines: dímer de timina. Un dímer de timina és una lesió greu del ADN i quan la ADN polimerasa se la troba s’atura.
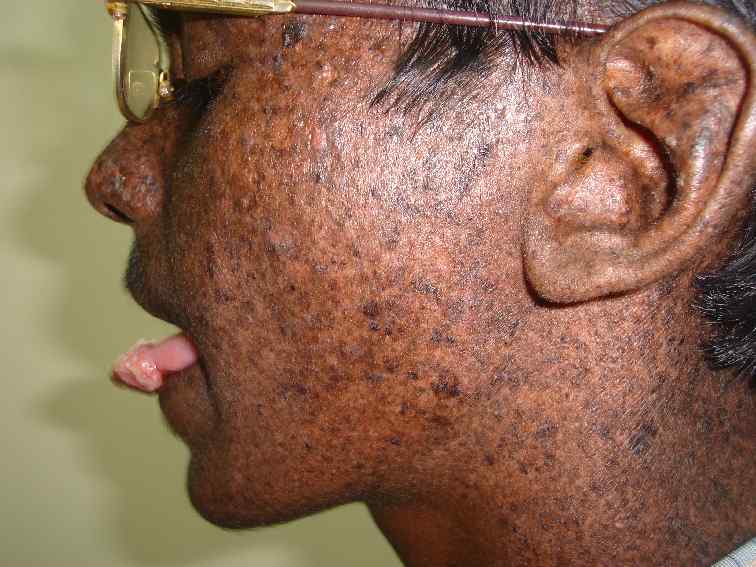
 La enfermetat Xemoderma picmentosum es produeix en nins on la quantitat de dímers de Timina és molt gran degut a que no tenen mecanismes de reparació del ADN. Són nins que han de quedar amagats a l’exposició solar ja que se les formen pigues que produeixen càncer.
La enfermetat Xemoderma picmentosum es produeix en nins on la quantitat de dímers de Timina és molt gran degut a que no tenen mecanismes de reparació del ADN. Són nins que han de quedar amagats a l’exposició solar ja que se les formen pigues que produeixen càncer.2. Radiacions ionitzants
Són radiacions electromagnétiques de longitud d’ona inferiors als UV i molt més energètiques. Són per exemple: raigs X, raigs gamma i les emisions de partícules radioactives: radiacions alfa i beta.
Provoquen la pèrdua d’electrons en alguns àtoms del ADN que queden en forma d’ions molt reactius.



Mutacions puntuals o gèniques
 1. Mutacions per substitució d´una base: canvis d´una base per una altra. Les substitucions provoquen l´alteració d´únic triplet , per tant si la mutació no dóna lloc a un triplet de STOP o no afecta a un aminoàcid del centre actiu del enzim, aquestes mutacions solen ser poc importants.
1. Mutacions per substitució d´una base: canvis d´una base per una altra. Les substitucions provoquen l´alteració d´únic triplet , per tant si la mutació no dóna lloc a un triplet de STOP o no afecta a un aminoàcid del centre actiu del enzim, aquestes mutacions solen ser poc importants.
- Transicions: substitució d’una purina per una altra purina (A-G). O de una pirimidina per una la pirimidina (T-C).
- Transversions: subtitució d´una purina per una pirimidina o al inrevés.

2. Mutacions per pèrdua o inserció de nucleòtids:
- Delecció o pèrdua d’un PB
- Addició d’un PB.
Aquestes corresponen al 80% de mutacions espontànees. La addició o delecció d’un PB produeix un desplaçament de la pauta de lectura, la qual cosa sol provocar canvis dràstics en la proteïna que s’ha de codificar.
Mutacions cromosòmiques
Són canvis en l’estructura interna del cromosomes.
Més que nous canvis del material genètic, impliquen reordenació del material ja existent.

Pe: delecció en el cromosoma 5 provoca el síndrome de cri du chat. Aquests nins afectats: fan uns renous semblants en es miulo d’un moix quan ploren, microcefalia, retràs mental i no arriben a adults.
- Duplicació: repetició d’un fragment de cromosoma. La rèplica pot col-locar-se en el mateix cromosoma o en un altre o fins i tot forma un cromosoma nou. Posiblement la duplicació fa menys mal que la delecció.
- Inversió: és el canvi de sentit d’un fragment en el cromosoma. Pot ser paracèntrica si no inclu el centròmer o pericèntrica si inclou el centròmer.
- Translocació: és el canvi de posició d’un segment de cromosoma. Quan els fragments de cromosoma no són homòlegs la translocació és recíproca.
Si la translocació es realitza entre fragments diferents s’anomena transposició. El síndrome de Down es pot produir degut a la translocació del cr 21 al 14.
Mutacions genòmiques
És una alteració en el nombre de cromosomes, que és propi de l’espècie.
Euploidia:
És una alteració en el nombre normal de dotacions haploides (n) (múltiple de n). De fet la poliploidia és l’existència de més de dues dotacions cromosòmiques: triploidíes 3n, tetraploidíes 4n ...
Sol ser poc freqüent en animals però més freqüent en plantes, donant lloc a fulles o fruits més grossos, la qual cosa és beneficiós per augmentar la producció.
La causa sol ser un error en la divisió de maduració de l'òvul o l'esperma.


Aneuplodia:
És l’alteració en el nombre normal de cada un dels tipus de cromosomes – normalment dues còpies de cada cromosoma -. Poden ser:
Nulosomies: no hi ha cap dels dos cromosomes 2n-2
Monosomies: només hi ha un cr del parell normal 2n-1
Trisomies, tetrasomies: 3, 4 còpies d'un mateix cr 2n+1, 2n+2
- Síndrome de Down: Trisomia del crosoma 21 (tenen 47 cr: 2n+1). Aquest és el més comú i més conegut desordre cromosòmic. Més o manco 1 de 800 nins neixen amb el síndrome i la freqüencia augmenta en mares amb més de 35 anys. El síndrome fou descrit clínicament per Langdon Down en 1866, però el seu origen fou desconegut durant quasi un segle. Des del punt de vista poblacional es van donar comte de què la freqüència augmentava en elevar la edat materna i la gran concordància entre bessons univitelins. No fou fins al 1959 que es va descobrir que tenien 47 cr. Tenen plec en els ulls, llengua gorga, coll curt, front pla, cara plana i ampla, trets facials orientals d’aquí el terme “mongolisme” que és totalment inapropiat. Baix desenvolupament intelectual, però no nul. La seva vida és curta, envelleixen ràpidament, amb arritmies cardíaques i pateixen la síndrome d’Alzeimer.


- Síndrome d’Edwards: Trisomia del cr 18 (2n+1). La incidència és de 1 cada 8000 nins. En general el 95% de concepcions de trisomia del cr 18 són abortamnets espontanis. Són persones amb retràs mental i en el desenvolupament. Hipertensió. Orelles deformades, coll curt i ulls petits.


- Síndrome de Turner: només té un cromosoma X (monosomia: 44+X). La incidencia és 1 cada 5000 nines. Presenten una estatura baixa, disgènesis gonadal, infantilisme gonadal, elevades anomalies renals i cardiovasculars. Els tractaments hormonals poden corregir algunes d´aquestes característiques, encara que l’estarilitat sol ser irreversible.

- Síndrome de Klinelfelter (47 cr, XXY): són al-lots aparentment normals fins a la pubertat, llavors apareixen signes d’hipogonadisme: testicles petits i característiques sexuals secundàries poc desenvolupades. La gran dosis de X fa que tenguin pit. No hi ha espermatogènesis. Freqüentment hi ha retràs mental.

- Trisomia del cr X: (47 cr, XXX): són la contrapartida del síndrome de Klinelfelter en la dona. Són dones amb mames poc desenvolupades i genitals externs infantils. Molts de casos queden sense diagnosticar.
- Síndrome de la insensibilitat al andrògen completa: conegut com feminització testicular. Les persones són cromosòmicament mascles 46XY, però amb una aparent genitalia femenina normal, amb una vagina cega i sense úter. Poseeixen testicles que poden estar en el abdomen. La incidència és de 1 cada 20000nins.

- Verificación de género en el deporte
Prevenció i diagnòstic de malalties genètiques
-
Diagnòstic prenatal
-
Prevenció primària
- En primer lloc es tracta de tenir controlat els mutàgens.
- Estudi de poblacions i de famílies: es tracta de conèixer els portadors i incidència. Si es coneixen els portadors es pòden fer estudis posteriors per evitar fills en deficiències.
- Consell genètic: proporcionar informació sobre el risc d'un individu o els seus descendents siguin afectats per una malaltia genètica. Es fa mitjançant l'elaboració d'una història clínica i d'un arbre genealògic.
-
Prevenció secundària
- Diagnòstic prenatal
Si no hi ha casos anteriors en la família es realitza un estudi prenatal a totes les dones embarassades.
- Analítiques:
1r trimestre
- Analítica de sang i orina:bioquímica, hemograma, rubeola, sífislis, toxoplasmosis, hepatitis B i C, VIH.
- Determinació del grup sanguini i del grup RH (test de Coombs).
- Triple screening: marcadors bioquímics (PAPP-A, Beta-HCG i fracció lliure)
2n trimestre
- Analítica bàsica de sang i orina: bioquímica i hemograma.
- Test de Coombs si la mare és Rh-.
- Hepatitis, Toxoplasma.
- Test O'Sullivan: risc de tenir diabetes gestacional.
3r trimestre- Analítica bàsica de sang i: bioquímica i hemograma
- Test de coagulació.
- Streptococco: si la mare es portadora un 50% de nounats neixen colonitzats i un 2% pot desenvolupar una infecció greu.
- Ecografies
1r Trimestre: (setmana 12) detectar alteracions estructurals congènites. (Traslucència nucal)
2n trimestre: (setmana 20) Morfològica que pot detectar malformacions en òrgans i estructures.
3r trimestre: (setmna 32-35) per conèixer l'estat de la placenta, la posició fetal o la quantitat de líquid amniòtic
- Amniocentesis: són anàlisis opcionals dutes a terme en la dona embarassada que ajuden a detectar trastorns genètics del fetus abans del naixement. Consisteix a extreure una mostra de líquid amniòtic de l'úter mitjançant una agulla que travessa l'úter. L'amniocentesi es realitza a partir de la 15a setmana aproximadament.
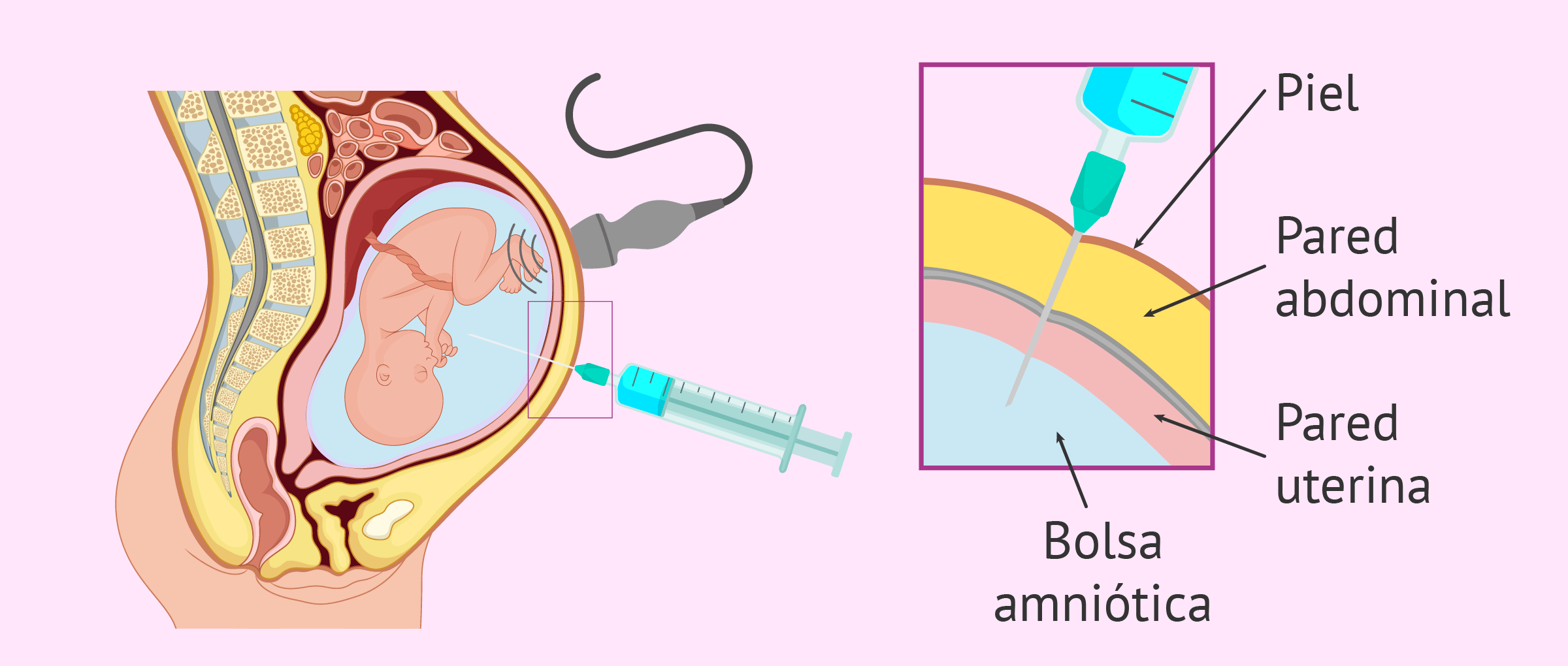
- Biopsia corial: La biòpsia o mostreig de les vellositats coriòniques (MVC en anglès), igual que l'amniocentesi, és una anàlisi opcional duta a terme en la dona embarassada que ajuden a detectar trastorns genètics del fetus abans del naixement. El MVC consisteix a extreure una mostra de placenta a través de la vagina emprant un catèter o agulla, que després serà analitzada al laboratori. Es realitza habitualment entre les setmanes 10 i 12 de l'embaràs.


- Cordocentesi: s’obtenen mostres de sang fetal, però la tècnica és didícil i té risc. El risc a malaltia ha de ser superior al perill que té el fetus amb aquesta prova.

-
Diagnòstic postnatal
- Prova del taló: s'extreu sang del nounat per detectar possibles trastors metabòlics relacionats amb la deficiència o manca d'un enzim
 - Fenilcetonúria: és una malaltia metabòlica congènita. Consisteix en l'acumulació de fenilalanina, la qual cosa provoca en la majoria dels casos retard mental. El tractament consisteix en realitzar una dieta restrictiva (llets especials i vegetariana) que, instaurada precoçment i ben controlada, aconsegueix que el bebè tengui un desenvolupament normal. Afecta a 1 de cada 10.000 bebès.
- Fenilcetonúria: és una malaltia metabòlica congènita. Consisteix en l'acumulació de fenilalanina, la qual cosa provoca en la majoria dels casos retard mental. El tractament consisteix en realitzar una dieta restrictiva (llets especials i vegetariana) que, instaurada precoçment i ben controlada, aconsegueix que el bebè tengui un desenvolupament normal. Afecta a 1 de cada 10.000 bebès.
- Hipotiroïdisme congènit: també és una malaltia metabòlica congènita. Consisteix en la manca de producció d'hormona tiroide en els primers mesos de vida. Provoca retard mental irrecuperable, però si s'inicia el tractament en els primers dies, el desenvolupament cerebral serà completament normal. Afecta a 1 de cada 3.000 bebès.
- Fibrosi quística:és una malaltia genètica les alteracions més freqüents de la qual són malaltia pulmonar crònica i insuficiència pancreàtica. La detecció de mutacions permet establir un diagnòstic precís. El tractament permet una millor qualitat de vida. Afecta a 1 de cada 2.500 bebès.
Biotecnologia i enginyeria genètica
-
Biotecnologia: utilització de sistemes biològics, éssers vius o derivats en la creació o modificació de productes o processos d'interès per a les perssones.
La biotecnologia fa molt temps que s'utilitza:
- Agricultura i ramederia: millora animal i vegetal, per obtenir organismes més resisteentsa plagues i malalties, més productives. Per selecció artificial.
- Indústria alimentària: microorganismes per a la fabricació del vi, el pa, el iogurt, el formatge,...
- Medicina: per produir antibiòtics o vacunes.
Enginyeria genètica
-
Enginyeria genètica: conjunt de tècniques de manipulació de gens al laboratori fetes amb un propòsit concret que sigui de profit per a les persones.
-
Tècniques d'enginyeria genètica:
- Enzims de restricció: proteïnes capaces de tallar l'ADN en punts específics.
Com funciona un enzim de restricció?
- ADN lligases: proteïnes que permeten unir fragments d'ADN de diferents.
- Vectors de transferència: molècules d'ADN que es poden reproduir autònomament i que serveix per transportar gens (plasmidis o virus desactivats).
Tècniques d'enginyeria genètica

Reacció en cadena de la polimerasa (PCR)

Tècnica de la PCR
PCR (Reacció en cadena de la polimerasa)
Kary Mullis (1986)![]()
Entrevista
Clonació


Projecte Genoma humà
El Projecte Genoma Humà (PGH) (Human Genome Project -HGP- en anglès) fou un esforç internacional de recerca per determinar la seqüència del genoma humà i identificar-ne els gens que conté. El Projecte, promogut i coordinat per dues agències governamentals dels Estats Units, va comptar amb contribucions des de diverses universitats estatunidenques i altres institucions públiques del Regne Unit, França, Alemanya, Japó i la Xina, que constituïren el Consorci International de la Seqüenciació del Genoma Humà (International Human Genome Sequencing Consortium -IHGSC- en anglès). El PGH s'inicià formalment el 1990 i es va completar el 2003, dos anys abans del previst.
Tècnicament consistia en determinar les posicions relatives dels nucleòtids (o parells de bases) de l'ADN humà i identificar els gens que hi són presents.

- Anglès
- Castellà
Encode project
Presentan el nuevo mapa del genoma humano.
Genome Data viewer
Per saber més i entendre millor
- Animals transgènics
Información bàsica - Introducción de genes (EIBE)
La oveja transgénica (EIBE)
El sumosalmón
- Plantes transgèniques
Introducción - Plantas transgénicas
Plantas transgénicas: problema y posibilidades
Casos reales
¿Qué plantas?
Les mutacions i l'enginyeria genètica.
L'origen i l'evolució de la vida
1. L'origen de la vida

L'origen de la vida ha estat des de l'antiguitat un tema d'interès per a pensadors, filòsofs i científics.
Els primers a proporcionar explicacions raonades basades en l'observació i no en fenòmens sobrenaturals o mitològics van ser els grecs. Aristòtil va exposar la idea de la generació espontània, els éssers vius sorgien, segons ell, d'altres éssers o objectes que en res tenien a veure amb l'existència d'uns progenitors semblants a ells; aquesta idea va prevaler fins en l'Edat mitjana i va perdurar fins i tot fins al segle XVIII .
Van Helmont assegurava en el segle XVII: «basta col·locar roba bruta en una bóta, que contingui a més uns pocs grans de blat, i al cap de 21 dies apareixeran ratolins».
Les primeres experiències científiques en contra d'aquesta teoria es van dur a terme per Redi (1668) i Spallanzani (1767), però fins a 1862 Louis Pasteur no va demostrar definitivament la seva falsedat.
Oparin en 1923 va proposar la hipòtesi de la «sopa primitiva», en la qual sosté que la vida es va originar en la Terra espontàniament per la influència de l'energia solar i descàrregues elèctriques sobre els gasos –amoníac, vapor d'aigua, metà, etc.–presents en l'atmosfera primitiva, que van poder reaccionar i generar molècules orgàniques senzilles com a aminoàcids simples. Aquestes molècules s'acumularien en els oceans formant el que ell va denominar «sopa primordial» on concentrades van poder interaccionar i evolucionar cap a coacervats i les primeres formes de vida.
Stanley Miller en 1953 va reproduir en el laboratori les condicions atmosfèriques primitives propostes per Oparin. En un circuit tancat va introduir una mescla d'aigua, amoníac, metà i hidrogen, va col·locar uns elèctrodes per a provocar descàrregues elèctriques sobre la mescla durant dies, al cap dels quals va poder comprovar la síntesi d'aminoàcids, molècules orgàniques que formen proteïnes.
Generación espontanea.
Atmosfera primitiva.
Estromatolitos, cianobacterias,...
Síntesis prebiótica: Miller.
El origen de la vida en la Tierra. (Documental complet. Canal Historia)
Origen de la vida: fuentes hidrotermales
Hallados los fósiles más antiguos de la Tierra (SINC - Marzo 2017)
2. Evolució Biològica

Espècie: conjunt d'individus que poden reproduir-se entre si i donar una descendència fèrtil (només es refereix a organismes amb reproducció sexual, no hi podem inclore els organismes amb reproducció asexual – moneres o organismes fòssils)

Teories no evolucionistes 
Linné (Carl von 1707-1778), pare de la nomenclatura binomial.


 Génesis 1:1-3:24 - El Principio
Génesis 1:1-3:24 - El PrincipioEn el principio Creó Dios los cielos y la tierra.
Y la tierra estaba sin orden y Vacía. Había tinieblas sobre la faz del océano, y el Espíritu de Dios se movía sobre la faz de las aguas.
Entonces dijo Dios:
Al primer día: “ Sea la luz; y fue la luz..”...
...
Al sexto: “Entonces dijo Dios: Hagamos al hombre a nuestra imagen, conforme a nuestra semejanza;...
I el séptimo día descansó.
Cuvier (George – 1769-1832). pare de l'anatomia comparada i de la paleontologia. Explica que els fòssils eren les restes d'organismes que havien existit en el passat però descartà que pogueren ser antecessors dels organismes actuals.


Diluvi Universal i l'Arca de Noé- Darrera catàstrofe?



Teories evolucionistes
- Tendència a la complexitat: l'evolució té lloc des de les espècies més senzilles i primitives fins a les més complexes que s'observen a l'actualitat.
- Aparició d'adaptacions al medi per ús i desús dels òrgans (la funció crea l'òrgan): variacions en el medi indueixen a canvis en els costums, es comencen a utilitzar uns òrgans que es desenvolupen i altres òrgans entren en desús per la qual cosa s'atrofien.
- Herència dels caràcters adquirits. Els canvis que tenen lloc en els organismes durant la seva per adaptar-se al medi es transmeten als descendents.
*Herència epigenètica: les condicions ambientals determinen que molècules s'associin a l'ADN i que enflueixin en la seva expressió, aquestes associacions són estables i poden passar als descendents.


- Darwin (Charles 1809-1882- obra l'Origen de les espècies 1859) i Wallace (Alfred Russell 1823-1913).

:max_bytes(150000):strip_icc()/HMS-Beagle-3000-3x2gty-579e23883df78c32765450a0.jpg)
- Malthus (Thomas 1766-1834): la producció d'aliments augmenta més lentament que el creixement de les poblacions humanes, aquest conflicte genera una lluita per l'existència.
- Lyell (Charles 1797-1875): geòleg que proposava que els processos geològics del passat són semblants als actuals i tenen lloc de forma lenta i gradual (Actualisme i uniformisme).
- L'alta capacitat reproductora: si el nombre d'individus d'una població no creix indefinidament és perquè els recursos alimentaris són limitats.
- Variabilitat de la descendència: s'observa que els organismes que es reprodueixen de manera sexual, els descendents presenten diferències encara que tinguin els mateixos pares (excepte bessons univitalins). Aquestes diferències s'acumulen generació rere generació i comporten: o la mort de l'individu, o no tenen cap tipus de rellevància, o suposen un avantatge davant un canvi en l'ambient.
- Selecció natural: la lluita per la supervivència que té lloc entre els individus quan hi ha poc aliment provoca que uns individus morin i altres sobrevisquin transmetet els seus caràcters a la generació següent.
- La variabilitat és heretable. Herència dels caràcters adquirits??
- Els descendents no són rèpliques dels progenitors, són combinacions.
- Els éssers vius fan tot el possible per sobreviure i tenir descendència.
- Selecció sexual: la varietat de formes, color i comportaments d'alguns animals només es pot explicar per aconseguir èxit reproductiu.
- Coevolució: moltes espècies interactuen tan estretament amb altres espècies que s'influeixen mútuament i evolucionen conjuntament


3. Proves de l'evolució
1. Proves taxonòmiques
Les espècies es poden agrupar per les seves semblances.
Apareixen els tàxons jeràrquic.Taxonomia: mètode per a classificar els éssers vius observant i analitzant les característqiues internes i externes, així es distribueixen en conjunts o subconjunts cada vegada més petits (Tàxons) d'acord amb les característiques que tenen en comú.
Domini Regne Embrancament Classe Ordre Família Gènere Espècie
El fet de poder agrupar espècies diferents és una prova de l'evolució i que pertant provenen d'un avantpassat comú.



Cas de l'ornitorrinc (ovípar amb bec corni – rèptils, pèl i succió de llet - mamífers)
Cas Archaeopteryx (dents i coa – rèptil, ales amb plomes – au)



2. Proves paleontològiques.
Estudi de restes fòssils.
Es poden aconseguir trobar sèries contínues de fòssils que permeten seguir l'evolució d'alguna espècie.
Exemple: sèrie evolutiva del cavall.

Estudi de l'anatomia comparada (comparar diferents òrgans del cos d'espècies diferents, en estructura i funció).
Així els òrgans poden ser:
- Òrgans homòlegs- Mateixa estructura però forma i funció diferent.
En alguns animals podem observar òrgans que tenen un aspecte diferent perquè tenen funcions distintes però que responen al mateix model d'estructura bàsica, aquests animals descendeixen d'un avantpassat comú.
Exemples d'evolució divergent.

Evolució convergent adaptativa - diferent origen - mateixa funció. Davant les mateixes necessitats, les adaptacions són similars.
Són proves de la nostra evolució - canvi.

.jpg)



4. Proves biogeogràfiques
Espècies que viuen en llocs allunyats són diferents però encara relacionades, conserven trets comuns perquè provenen d'un avantpassat comú. (Relacionat amb l'especiació simpàtrica)
Exemples:
- Moneia (Amèrica) i ximpanzè (Àfrica)
- Elefants africà i indi.
- Austràlia marsupials.


Estudi del desenvolupament embrionari.
Les primeres etapes del desenvolupament embrionari mostren moltes semblances, i més com més pròxims són els animals.
Llei de la biogenètica: l'ontogènia recapitula la filogènia. (Haeckel 1834-1919, Llei biogenètica)
El procés de desenvolupament embrionari (ontogènia) és un repàs breu de l'història evolutiva (filogènia) de l'espècie a la qual pertany l'embrió. Segons aquesta llei, durant les primeres fases del desenvolupament embrionari d'una espècie, es manifesten algunes característqiues que tingueren les espècies ancestrals, encar que posteriorment desapareixen.
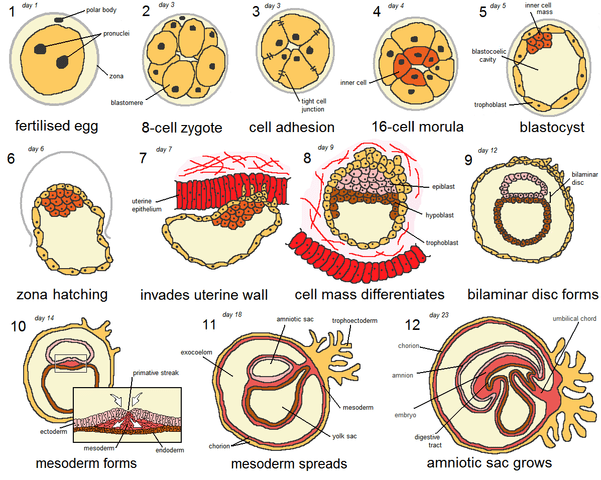
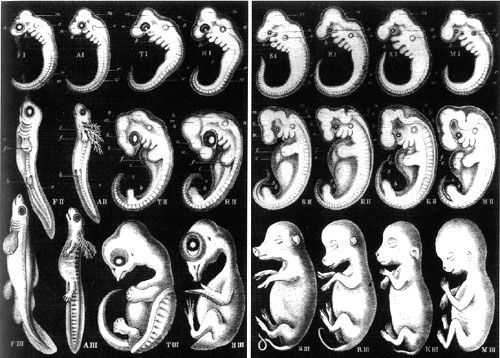

6. Proves bioquímiques o de biologia molecular.
Proves bioquímiques o de biologia molecular.
Estudi d'algunes molècules presents en els éssers vius.
Hi ha una gran semblança entre molècules i més com més pròxims són els éssers vius.
- Proves basades en proteïnes:
- Proves basades en els àcids nucleics:
S'obtenen comparant les seqüències de nucleòtids de l'ADN i de l'ARN entre espècies. En els estudis filogenètics, per evitar la variabilitat que existeix en l'ADN, es treballa amb l'ADN mitocondrial, amb l'ADN del cromosoma Y o amb els gens que codifiquen l'ARN ribosòmic.
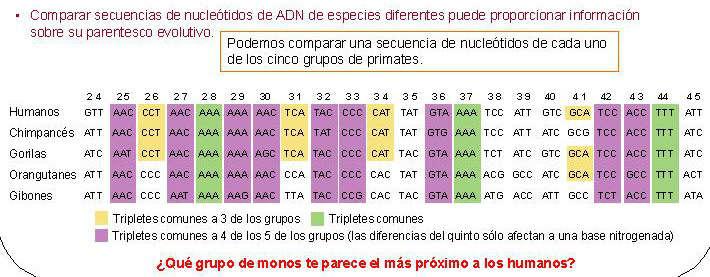
- Proves serològiques o immunològiques:
Són les basades en l'estudi comparat de les reaccions d'aglutinació de la sang en els diferents organismes.
S'introdueix sang d'un individu d'una espècie en un altre d'una espècie diferent, a fi que aquest últim fabriqui anticossos específics contra les molècules (antígens) de la sang rebuda. Després, si aquesta sang carregada d'anticossos es posa en contacte amb sang de l'espècie donant, hi ha un grau d'aglutinació; si es posa en contacte amb una espècie semblant a la del donant, el grau d'aglutinació és més elevat; i si es posa en contacte amb la sang d'una espècie diferent, el grau d'aglutinació és molt baix. Aquests resultats es consideren una prova de l'evolució de les espècies.
Models evolutius actuals
- El procés evolutiu es basa en els principis darwinistes de la variabilitat de la descendència i de la selecció natural.
- La variabilitat de la descendència es deu a les mutacions (a l'atzar) i a la recombinació que té lloc durant la meiosi (noves combinacions de genotips).
- El que evolucionen són les poblacions no els individus. Un individu neix i mor amb el mateix genotip però les poblacions canvien a mesura que varien les freqüències gèniques (% de cada un dels al·lels que hi ha per a cada caràcter)
- Els factors que fan variar les freqüències gèniques són: selecció natural, les mutacions, les migracions i la deriva genètica.
- Una població donarà lloc a una espècie nova si hi ha aïllament reproductiu respectes a les altres poblacions.

Teoria neutralista de l'evolució molecular.
Kimura va observar que hi havia diferències d'aminoàcids d'una mateixa proteïna entre individus d'una mateixa espècie, per tant la major part de mutacions no són favorables ni desfavorables; són mutacions neutres (no són afectades per la selecció natural).
Les mutacions neutres poden introduir canvis perdurables en l'ADN molt ràpidament.
Si es coneix el nombre de substitucions de nucleòtids per unitat de temps en uns determinats gens i es considera que aquest ritme es manté costant, és posible calcular en quin moment va tenir lloc un determinat fet evolutiu.


Kimura. Comparació de 73 seqüències d'aminoàcids de la proteïna NP
- Les espècies romanen invariables o sense canvis importants durant llarg períodes de temps coneguts com a estasis i després i altres etapes de transformació molt ràpida o especiació.
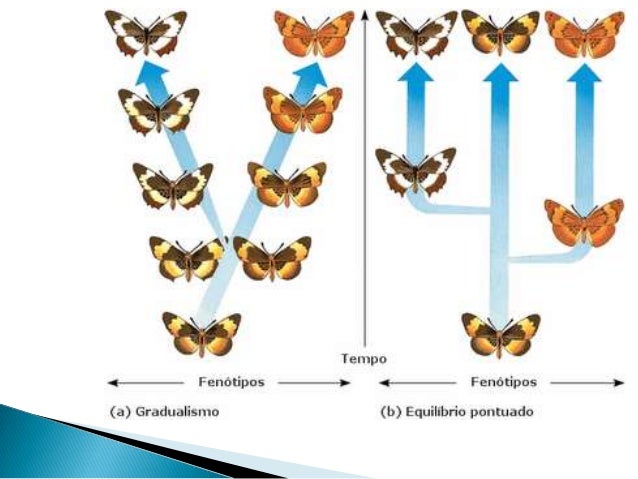
- Simbiogènesis. Lynn Margulis (1967)
Lynn Margulis va proposar la teoria de l'endosimbiosi que postula l'origen de la cèl·lula eucariota a través s'una sèrie de simbiosi entre organismes unicel·lulars de diferents espècies. Està acceptat l'origen bacterià dels mitocondris i cloroplasts.
Principis:
- El principal mecanisme d'obtenció de gens nous en una població són la transferència de gens i fins i tot genomes sencers, entre espècies (I no les mutacions)
- Defensa processos ràpids de formació d'espècies mitjançant l'establiment de relacions simbiòntiques entre ells (líquens: alga + fong)
4. Adaptació i especiació
Especiació: procés evolutiu de formació d'espècies (Evolució —►formació de noves espècies)
Espècie: dos individus són de la mateixa espèecie si a més de tenir trets físics semblants, es poden resproduir i la seva descendència és fèrtil.
Mecanismes d 'especiació - aïllament reproductor
Barreres geogràfiques: mars, rius, muntanyes, carreteres
2. Especiació simpàtrica - aïllament reproductor
- Temporal: mateix territori, però arriben al zel o a la floració en moments diferents.
- Etològic: comportament diferent que impedeix el festeig que dóna lloc a la còpula.
- Mecànic: diferència de mida, d'estructura dels òrgans copuladors.
- Gamètic: no hi ha atracció entre els gàmetes o el gàmete no sobreviu
3. Mutacions genòmiques
- Aneuploidia: alteració en el nombre normal de cada un dels tipus de cromosomes.
5. Mecanismes evolutius
- Mutacions
Les freqüències de mutació, per cèl·lula i generació, són molt baixes (1 mutació per cada 100.000 voltes que es copia el gen).
Les mutacions originen canvis que poden ser favorables o desfavorables, creen variabilitat.
- Selecció natural
Augmenta la proporció dels caràcters que aporten uns avantatges als individus ja que tenen més probabilitat de reproduir-se i de transmetre aquest caràcter als seus descendents.
- Deriva genètica
Un grup d'individus a l'atzar queda aïllat de la resta d'individus de la població i forma una població nova (Mecanismes d'especiació)
- Coll de botella: pocs individus sobreviuen a un esdeveniment que provoca molta mortalitat. (Lleones del Serengueti)


- Flux gènic- migracions
Les migracions d'individus d'una població a una altra poden alterar les proporcions gèniques.
La població originària, que migra, perd variabilitat i els seus gens es perden.
La població d'acollida augmenta la variabilitat, els individus nous hi aporten el seus gens.

6. Hominització
Adaptacions:
- Permet visualitzar l'horitzó per damunt la vegetació.
- Marxa més lenta però menys costosa i permet recórrer grans distàncies.
- S'estreny el canal pelvià i el part és més complex i dolorós.
- Augment de mida del polze, que permet agafar i dur a terme feines de precisió.
- Augment de la capacitat cranial.
- Aplatament de la cara, que el fa més lleuger.
El viaje de la evolución. (El joven Darwin)

Saber més per aprendre més
 Darwin. El padre de la evolución.
Darwin. El padre de la evolución.- Darwin
- El viaje del Beagle
- La evolución i la evolución del hombre
- Ciencia vs religión
- Hablan los expertos
Charles Darwin
Alfred Russel Wallace

El joc de l'evolució (Biston betularia): fes de depredador i veuràs com actua la selecció.
http://todosobrelaevolucion.org.mx/index.html
Estructura de la Terra
1. Origen del Sistema Solar.
Hipòtesi de l'acreció planetesimal.
2. Origen de la Terra.
Model de capes:
- Geosfera: Fa 4.500 Ma
Gran bolla de roca fosa, calenta per:
- Impactes de planetesimals durant l'acreció. (El xoc d'asteroides durant els primers milions d'anys de la formació de la Terra va fer que augmentés el seu tamany i la temperatura.)
- Desintegració d'isòtops radioactius (K-40 o U-235). Les roques de l'interior de la Terra contenen urani, plutoni o tori que quan es desintegren alliberen una gran quantitat d'energia que es transmet per radiació.
- Gran esdeveniment tèrmic (GET) per diferenciació dels materials en capes de densiat creixent a causa de la gravetat.
(Els materials metàl·lics (majoritàriament ferro) es van enfonsar i el fregament va produir molta calor.)
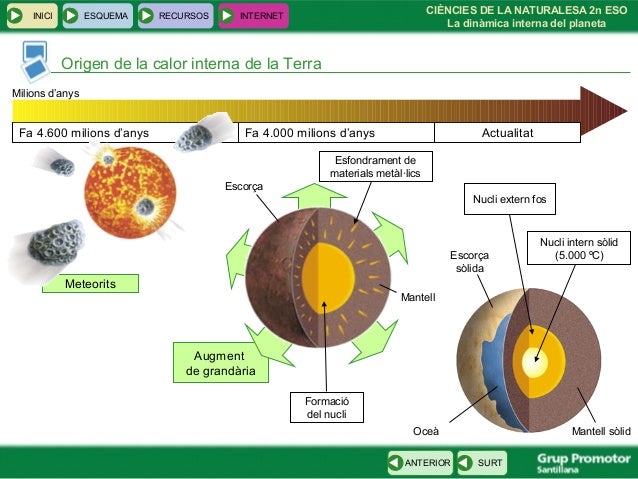

- Atmosfera i hidrosfera
- Durant el GET es van desprendre una gran quantitat de gasos.
- Posteriorment, l'aigua de l'atmosfera es va condensar.
- Biosfera
- La distància al Sol i les condicions fisicoquímiques, com la temperatura i l'existència d'aigua líquida, han afavorit la presència de la vida a la Terra.
3. El sistema Terra
3.2 Model estàtic o geoquímic
Capes geoquímiques:
Escorça oceànica
Zons de transició
Mantell inferior
3.3 Model dinàmic o geodinàmic
Capes geodinàmiques:
Litosfera continental
- Mesosfera: capa sòlida, encara que és capaç de fluir molt lentament. Permet el descens de les plaques litosfèriques i ascens deels plomalls de magma procedents de la zona D".
- Capa D": acumula calor procedent del nucli extern i n'escapen de forma episòdica. Els anomenats plomalls tèrmics ,columnes de magma molt calent que arriben a la litosfera i la perforen, i que originen els punts calents amb una activitat volcànica intensa.
- Endosfera:
Aquests corrents de convecció són els causants del camp magnètic terrestre.
4. L'estudi de l'estructura interna de la Terra
4.1. Mètodes directes
Sondejos, mines.


4.2. Mètodes indirectes
4.2.1. Meteorits
4.2.2. Les ones sísmiques
Sismes o terratrèmols: tremolors de terra causats per sacsejades brusques de l'escorça terrestre que són provocades pel desplaçament sobtat de grans masses rocoses situades a zones de fractura o falles.
Hipocentre: focus a partir del qual e spropaguen les vibracions en forma d'ones sśimiques.
Epicentre: punt de la superfície terrestre on arriben les ones sísmiques.
Els terratrèmols s'avaluen per la seva intensitat o la seva magnitud amb dos tipus d'escales:
- Escala de Mercalli, MSK (Medvedev - Sponheuer - Karnik) o EMS (European Macroseismic Scale):
Avalua la intensitat o sigui pels seus efectes destructius que produeix.
- Escala Richter:
Avalua la magnitud o sigui l'energia alliberada en un terratrèmol.
Tipus d'ones:
- Ones primàries P:
Ràpides.
Moviments de compressió i descompressió.
Són capaces de travessar materials sòlids com líquids, encara que redueixen la seva velocitat quan travessen els líquids.
- Ones secundàries S:
Més lentes que les ones P.
Ones perpendiculars a la trajectòria de l'ona.
Només es propaguen a través de materials sòlids.
- Ones superficials
Causen efectes catastròfics.
4.2.3. Magnetisme
4.2.4. Gravimetria
4.2.5. Isostàsia

Isostasia: equlibri de flotació de la litosfera rígida per damunt el mantell.
Subsidència: l'increment del pes sobre la litosfera pot provocar-ne l'enfonsament.
Sòn desplaçaments verticals de la litosfera, la qual tendeix a enfonsar-se en alguns llocs i a alçar-se en altres.
La litosfera s'enfonsa quan sobre es posen materials, per exemple: casquet de gel de milers de metres de gruix o sediments a una conca sedimentària (com un vaixell quan s'hi col·loca damunt tendeix a submergir-se)
La litosfera tendeix a alçar-se quan s'elimina part de la seva massa, per exemple: la desaparició d'una glaciació o l'erosió.
4.2.6. Flux tèrmic
Flux tèrmic: quantitat d'energia calorífica que arriba a la superfície terrestre des de l'interior del planeta. El calor es pot transmetre per conducció.
Gradient geotèrmic: augment de temperatura cap a l'interior de la Terra. A l'escorça té un valor mitjà de 3ºC/100m.
- En els primers quilòmetres la temperatura augmenta molt aviat, 30ºC per cada Km de profunditat
- En arribar als 300 km de profunditat les roques es troben a més de 1000 ºC de temperatura.
- En el centre de la Terra s'assoleixen temperatures de 5000ºC.

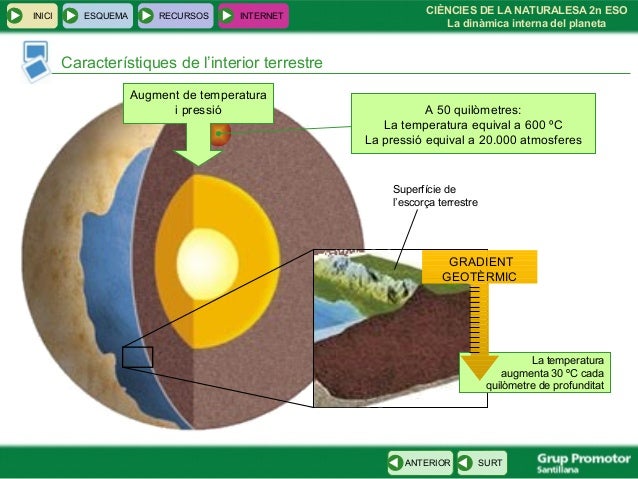
Estructura de la Terra
![]() Ese pequeño punto azul pálido (Carl Sagan)
Ese pequeño punto azul pálido (Carl Sagan)
![]() El agua de la Tierra és más antigua que el Sol.
El agua de la Tierra és más antigua que el Sol. ![]()
Contesta a les següents preguntes:
- Cerca que és l'isòtop deuteri.
- Quina finalitat presegueix l'experiment de L. Ilsidore Cleeves de la Universitat de Michiga?
- Explica amb les teves paraules l'experiment anterior.
- A quina conclusió s'ha pogut arribar?
3. Quants sísmografs hi ha a la Terra? Xarxa de sismógrafs internacional.
5. Vols saber més.... visita IRIS.
Dinàmica de la Terra. Tectònica i relleu.
1. Model dinàmic de la Terra.
2. Tectònica de plaques.
3. Proves de la Tectònica de plaques.
3.1. Derica continental de Wegener
 - Proves geogràfiques
- Proves geogràfiques
- Proves paleontològiques
- Proves geològiques
- Proves paleoclimàtiques
3.2. Paleomagnetisme terrestre. Edat del fons oceànic.
3.3. Distribució geogràfica dels fenòmens sísmics i volcànics.
4. Teoria de Tectònica de plaques.
4.2. Límits de les plaques litosfèriques
Vall del rift, llacs i mar lineal
Obducció
- Fenòmens intraplaca
5. Cicle de les roques

- Roques magmàtiques: s'originen pel refredament o consolidació d'un magma.
Granit
- Roques volcàniques: refredament ràpid, els minerals formen cristalls no observables a simple vista o no són cristalls.
Basalt
Traquita
Escòria volcànica
Pumicita - o pedra tosca
Esquist
Gneis
Quarsita
Marbre
Sediments ——►(procés de diagènesi: compactació i litificació) ——►Roca sedimentària
Es classifiquen pel tamany de gra i pels minerals que el formen:
Arenisques - marés
Calcària
6. El relleu de la superfície terrestre
Si tallassim longitudinalment la Terra podriem veure que l'escorça oceànica i continental no són continues sinó que hi ha una gran diversitat de formes, formes del relleu.

Escorça continental:
- Grans planes o escuts (6): grans extensions pràcticament horitzontals (plana de l'Amazones - Amèrica del Sud, Sàhara - Àfrica)
- Plataformes continentals (8): comprenen des de la línia de costa fins a uns quants quilòmetres mar endins. Són la vorera dels continents. Profunditat màxima de 300m. La plataforma continental acaba en una gran davallada fins a arribar a l'escorça oceànica, aquest fort pendent s'anomena talús continental (9).
Escorça oceànica:
- Serralades o dorsals oceàniques (3): zona elevada de les conques oceàniques. Les dorsals s'interconnecten per tots els oceans de la Terra i formen un conjunt de més de 70.000 Km de longitud.
- Fosses oceàniques (10): són les zones més profundes dels oceans. On es posen en contacte l'escorça oceànica i la continental. La fossa més profunda és la de les Mariannes amb 11.034 m de profunditat.
- Planes abissals (4): les zones planes més extenses del planeta, situades a una profunditat mitjana de 4.000 a 4.500 m
- Volcans submarins (5): relleus aïllats enormes, que en alguns casos poden arribar a la superfície de l'oceà i originar arxipèlags volcànics (Hawaii).
6.1. Plegaments
Plecs: deformacions plàstiques de les roques produïdes per forces de compressió durant molt de temps.
Parts
 - Xarnera: zona de major curvatura del plec.
- Xarnera: zona de major curvatura del plec.
- Línia de xarnera o eix de plec: línia que uneix els punts de major curvatura d’una superfície del plec..
- Plànol axial: plànol que conté totes les línies de xarnera i talla el plec.
- Nucli: part més comprimida i més interna del plec.
- Flancs: meitats en què divideix el plànol axial a un plec.
- Capbussament: angle que forma l’eix de plec amb una línia horitzontal continguda en el pla axial.
Tipus
 Segons la posició de les capes
Segons la posició de les capes
- Anticlinals
- Sinclinals
- Recte o simètric
- Inclinat
- Colze
- Tombat o ajagut
6.2 Falles
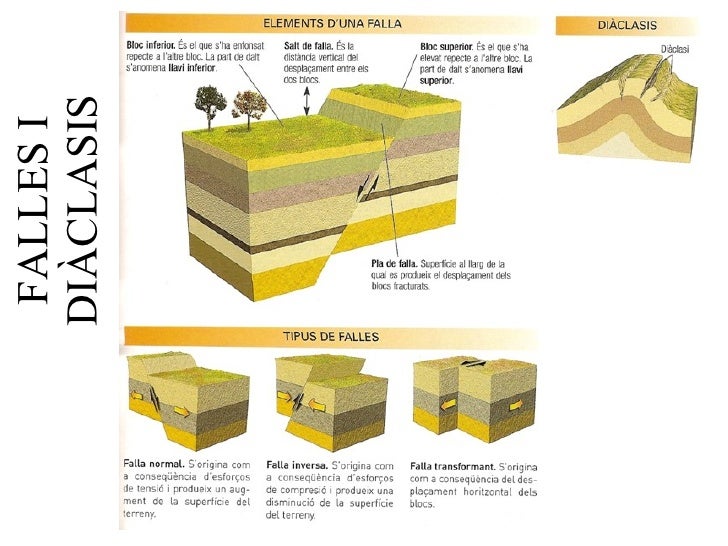
6.3 Vulcanisme
En general en el mantell les temperatures són més elevades però les altes pressions mantenen les roques en estat sòlid. En zones del mantell més calentes o menys pressió les roqes poden iniciar la fusió i formar magma.
El magma són mescles de roques foses, minerals cristal·litzats i gasos.
El magma és menys dens que les roques sòlides del seu voltant i tendeixen a pujar. Si aconsegueixen sortir per una factura de l'escorça es produiex una erupció volcànica.
En una erupció volcànica el magma perd els gasos i surt en forma de lava.
Productes volcànics.
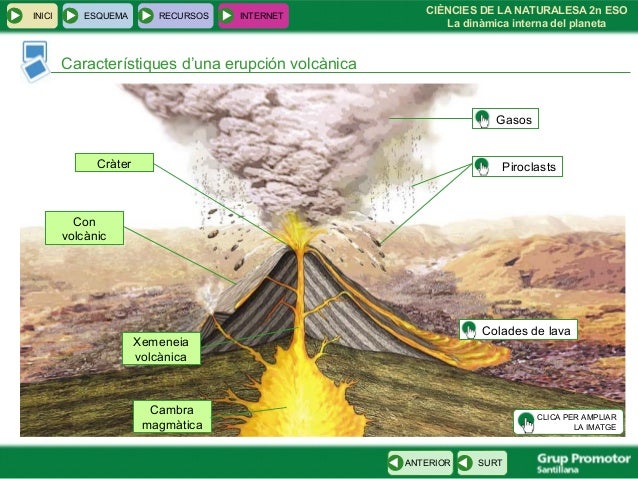
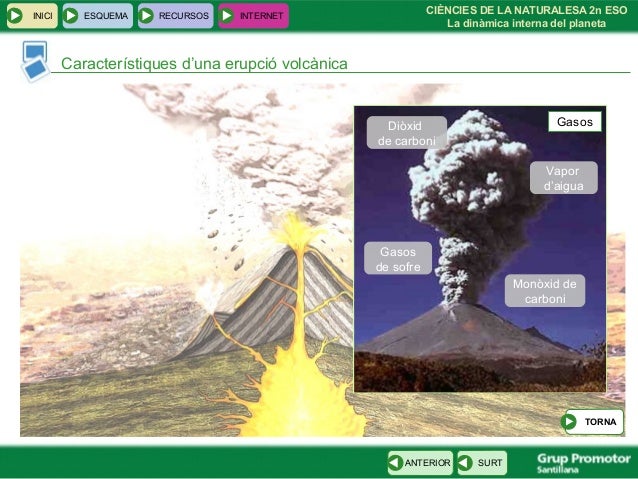
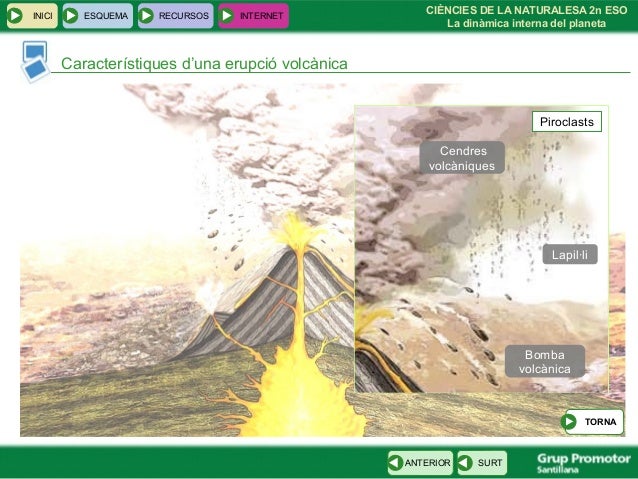 En una erupció volcànica s'expulsen materials en els tres estats:
En una erupció volcànica s'expulsen materials en els tres estats:- Gasos: diòxid de carboni CO2, vapor d'aigua H2O, monòxid de carboni CO, gasos de sofre
Magma fluïd menys perillós.
Magma viscós més perillós.
Segons el tipus d'edifici volcànic i le seva perillositat els volcans poden ser:
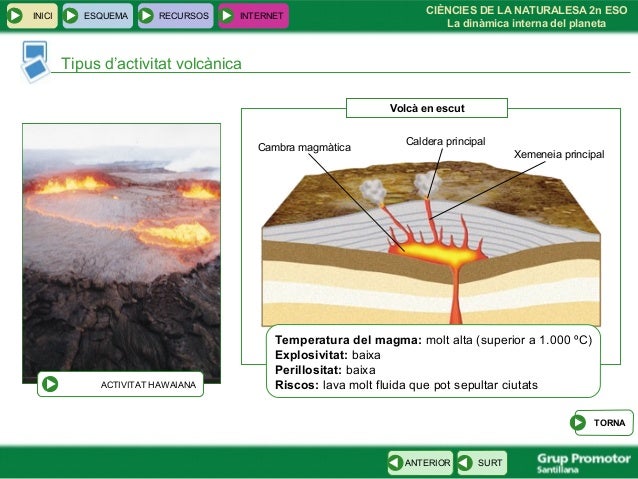 - Hawaians o en escut:
- Hawaians o en escut:Materials emesos: lava molt fluïda amb pocs piroclats
Edifici volcànic: escut - més extens que alt
Magma: temperatura molt alta
Explosivitat i perillositat: baixa
Riscs: la lava pot sepultar ciutats
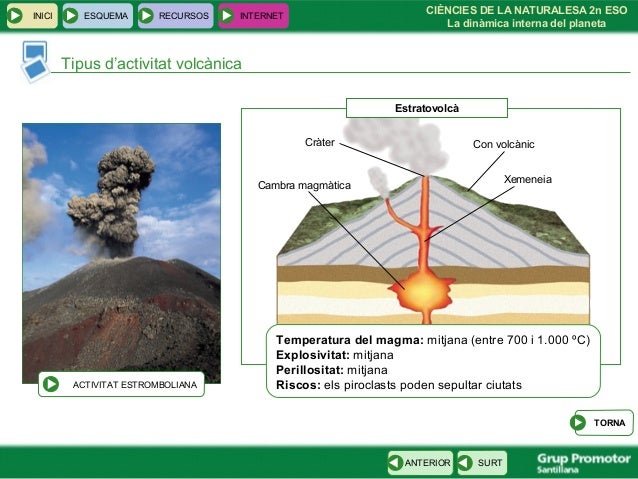 - Estrombolians:
- Estrombolians:Materials emesos: lava viscosa amb abundants piroclasts de totes les mides
Edifici volcànic: estratovolcà, més alt que extens (forma de con)
Magma: temperatura mitjana
Explosivitat i perillositat: mitjana
Riscs: els priroclasts poden sepultar ciutats
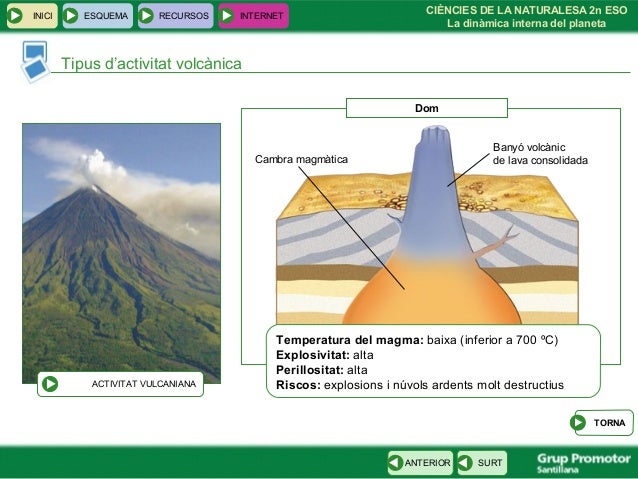 - Plinians:
- Plinians:Materials emesos: cendres mesclades amb gasos incandescents i abundants piroclasts.
Edifici volcànic: estratovolcà, més alt que extens (forma de con), la lava sol tapar el cràter i forma una cúpula anomenada dom.
Magma: temperatura baixa
Explosivitat i perillositat: alta
Riscs: els núvols ardents i les explosions són molt destructius.
La història de la Terra
1. L'edat de la Terra
James Hutton en el llibre "Teoria de la Terra" i pare de la geologia
Charles Lyell en els "principis de Geologia"
"El present és la clau del passat": els processos que havien donat forma a la Terra al llarg del temps eren els mateixos que encara ara es produeixen, com l'erosió, el vulcanisme o la formació de relleus per plegament.
"Els processos geològics actuen gradualment i durant llargs períodes de temps.
Neocatastrofista
Reconeix la coexistència de processos geològics d'intensitat i durada diferents. Hi ha processos graduals (erosió, formació d'un sòl) i alguns canvis catastròfics (meteorits, terratrèmols, tsunamis, erupcions volcànics,...)

2. Fòssils
Poden ser restes corporals, motlos, empremtes, restes biològics, ous, nius,...
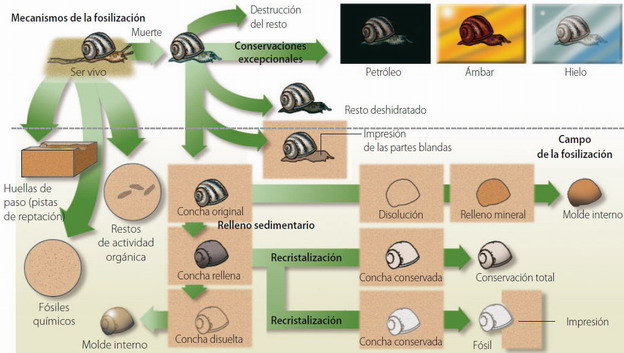
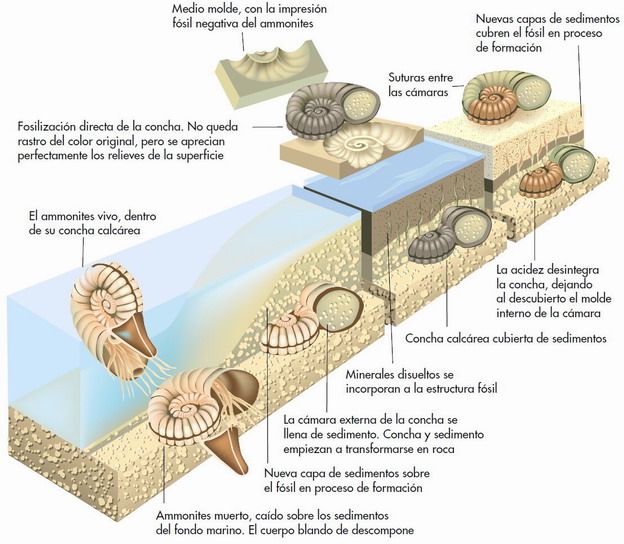
Són fòssils que aporten una informació molt valuosa sobre el substrat rocós que els conté.
Un fòssils per ser fòssil guia ha de complir les següents condicions:
- El procés de fossilització ha de ser fàcil.
- Ha de ser fàcil de reconèixer.
- Ha de tenir una dispersió geogràfica molt àmplia perquè se'n puguin trobar en llocs molt diferents.
- Han de ser molt abundants.
- Han d'haver viscut durant un període de temps curt.
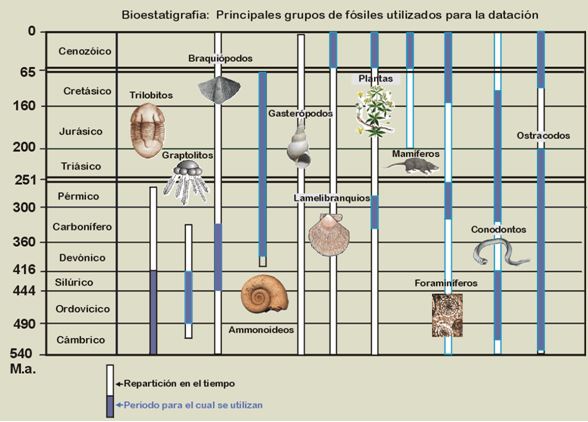
3. El temps en Geologia
Es poden diferenciar dos tipus de datació:
• Datació absoluta: l'edat d'un esdeveniment expressada en anys (sempre hi ha un marge d'error, podent ésser de milions d'anys, en funció del mètode que s'utilitzi).
- Varves glacials




Es base en l'estudi dels estrats.
Estrats: els materials sedimentaris es dipositen en capes anomenades estrat.
Els plans que limiten un estrat s’anomenen superfícies d’estratificació: la superior s’anomena sostre i l’inferior mur.
Els estrats s’agrupen en sèries estratigràfiques que abracen un determinat període de temps. El conjunt de totes les sèries s’integren en una sèrie estratigràfica total que comprèn la història geològica completa de la Terra.
Els estrats poden ser:
- concordants si s'han depositan de manera contínua.
- discordants sèrie sotmesa a plegaments, processos d'erosió i posterior dipòsit d'una altra sèrie d'estrats.
- Principi d'horitzontalitat original: el sediments que es depositen en una conca sedimentària s'adptan a la forma de la conca de manera horitzontal, aquesta disposició pot canviar per processos tectònics.
- Principi de continuïtat lateral: un estrat té la mateixa edat en tots els seus punts.
- Principi de successió faunística: els estrats tenen l'edat del fòssils que hi podem trobar.
- Pricipi de correlació: si una determinada associació de fòssils en un estrat i un lloc determinat el podem trobar en un altre lloc, els dos estrats tenen la mateixa edat.
- Principi de relació de tall: si una estructura geològica 1 (falla, plec o magma) retalla una altra estructura 2, la 1 s'ha produïnt fa menys temps.
- Principi de parsimònia (navalla d'Ockman): l'explicació més ssimple i suficient és la més probable, encara que no necessàriament vertadera.

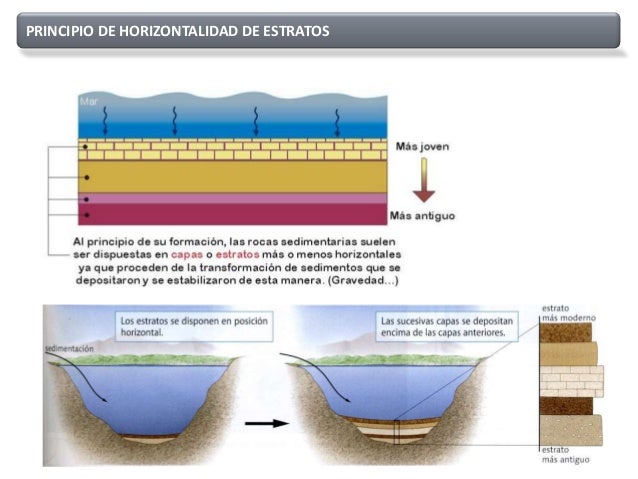

Estructura i dinàmica dels ecosistemes
1. La biosfera, l'ecosfera i els ecosistemes
La branca de la biologia o ciència que estudia la composició i funcionament dels ecosistemes és l'ecologia.
Biosfera: el conjunt de tots els éssers vius que habiten la Terra.


2. Biòtop
Per descriure'l hem de fer una llista del factors abiòtics que hi podem trobar.
Factors abiòtics: són les variables fisicoquímiques del medi que influeixen en la vida dels organismes.
Exemples de factors abiòtics:
Moviment de l'aigua: llacs, torrents, rius.
Temperatura: temperatura mitjana, temepratura màxima, temperatura mínima.
Precipitacions: quantitat de precipitacions i règim de precipitacions (igual en tots els mesos, concentrades en uns quans mesos,...)
Humitat relativa.
Vent.
Quantitat de llum.
Presència d'aigua.
Tipus d'aigua: salada, dolça.
Concentració d'oxígen i de diòxid de carboni.
Presència de contaminants
3. Relacions entre el biòtop i la biocenosi
La biocenosi provoca modificacions sobre el biòtop.
Els factors abiòtics influeixen en el tipus d'ecosistema, però també la biocenosi provoca modificacions sobre el biòtop amb el pas del temps.
Per exemple l'evolució del sòl:
Roca —► Líquens, plantes herbàcies —► Sòl —► Arbusts + sòl —► Arbres i arbusts + sòl
1. Els fenòmens atmosfèrics actuen sobre les roques de la superfície i en provoquen la meteorització.
2. S'hi instal·len éssers vius, que col·lboren a disgregar les rouqes i en provoquen compostos orgànics. En zones on lestemperatures a l'estiu són altes i les precipitacions escasses, es pot establir una biocenosi de líquens i plantes herbàcies. Després de 10 o 15 anys, la biocenosi provoca canvis en el biòtop: les arrels de les plantes van produint la disgregació de la part superficial de la roca calcària i s'allibera argila que conserva l'humitat.
3. El resultat és la formació d'una capa superficial que recobreix les roques i la formació d'horitzons. Després de 10 o 20 anys més, la creixent humitat del sòl i una capa més grruixada d'argila permeten el creixement d'alguns arbres.
4. El sòl madur permet una vegetació de port més gran i la instal·lació d'animals. Cada vegada el sòl és més argilós i méss humit, comença a desenvolupar-se una biocenosi diferent, hi ha més arbres i més grans i les cadenes tròfiques es transformen en xarxes tofiques de cada vegada més complexes. La relació entre biocenosi i biòtop s'intensifica i les relacions entre la biocenosi ees fan més estables.
4. Biocenosi
La biocenosi és el conjunt de tots els éssers vius que viuen en un ecosistema.
- El conjunt de poblacions que viuen a la mateixa àrea formen una comunitat.
Hàbitat: és el lloc físic d'un ecosistema on viuen els individus d'una espècie. L'hàbitat d'un ésser viu té les condicions naturals necessàries que li permeten viure-hi, si no és així canviarà d'hàbitat.
Nínxol ecològic: és el paper que du a terme o com actua una espècie en un ecosistema. P.e: com es comporta, de què s'alimenta, com es repodueix o com afecta a altres espècies del mateix ecosistema.
Si dues espècies tenen el mateix nínxol una d'elles tendirà a desaparèixer.
Girafes i zebres Sabana Girafes s'alimenten de les fulles de les branques altes
Zebres s'alimenten de les fulles de les branques baixes
Mussol i falcó Praderies o bosc Mussol s'alimenta de ratolins que caça de nit
Falcó s'alimenta de ratolins que caça de dia


El nínxol ecològic d'un organisme és el resultat de l'adaptació a l'ambient en què viu.
Anem a veure exemples d'adaptacions dels éssers vius als diferents medis:
Adaptacions a ecosistemes terrestres:
- Humitat:
- Ambients humits —► fulles grosses
Ambients secs —► fulles petites i estretes
- Ambients humits —► arrels pertites
Ambients secs —► arrels fondes i extenses
- Tenir estructures per emmagatzemar aigua —► cactus amb espines
- Secs —► animals amb closques i cobertes impermeables
Fulles amples com antenes
- Animals —► actius durant el dia per alimentar-se o per encalentir-se
- Homeoterms capaços de regular la seva temperatura corporal —► greix, pèls i plomes, suor, tremolor
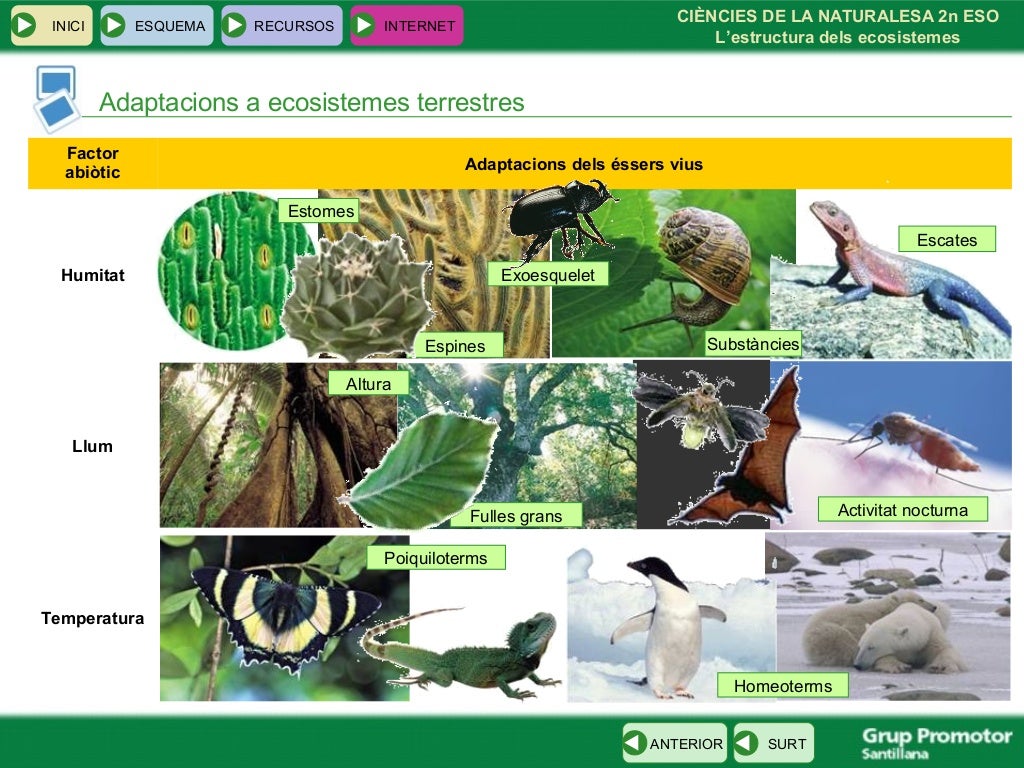
Adaptacions a ecosistemes aquàtics:
- Llum:
- Les algues tenen diferents pigments per poder captar la llum a diferents profunditats.
els animals tene formes arrodonides, es refugien o s'enterren.
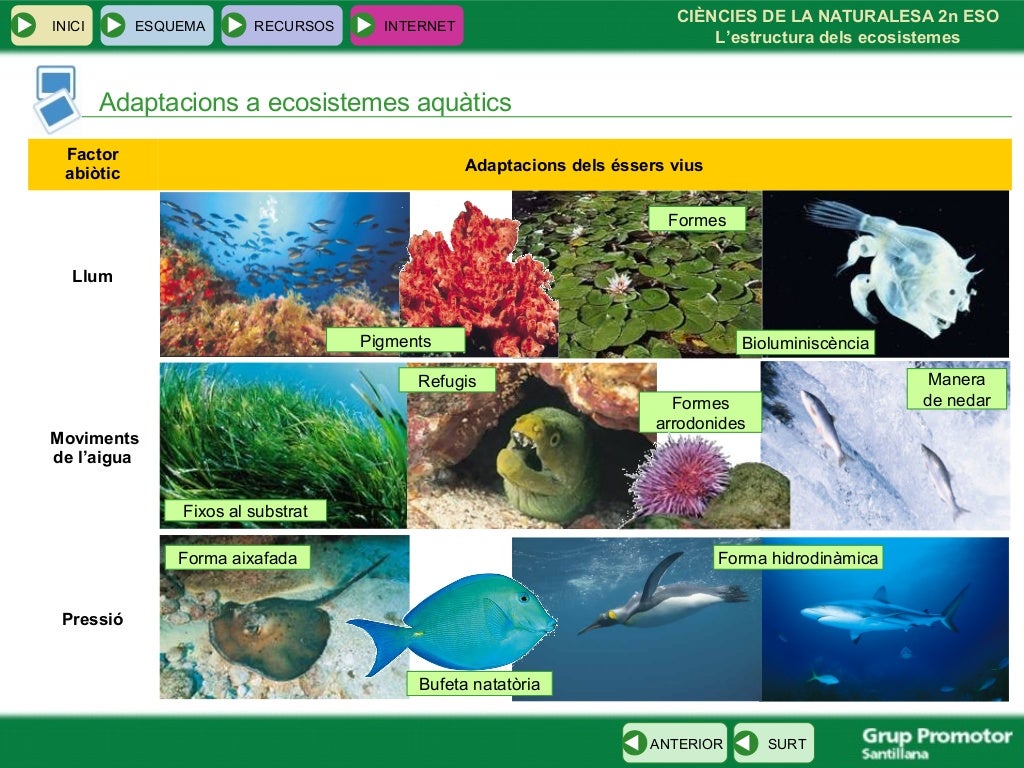
5. L'alimentació dels éssers vius en els ecosistemes
Aliments —► Nutrients —► Energia per viure
Els éssers vius es poden classificar segons la manera com obtenen aquests aliments en:
- Productors
- Consumidors
- Descomponedors
5.1 Productors
Tenen nutrició autòtrofa. Els productors no ingereixen matèria orgànica sinó que ells mateixos la fabriquen a partir de substàncies inorgàniques mitjançant un procés anomenat fotosíntesi.
La matèria orgànica que fabriquen és utilitzada pels consumidors.
Sol
Diòxid de carboni + Aigua + Sals minerals — — — — —►Glucosa (matèria orgànica) + Oxígen
5.2 Consumidors
Tenen nutrició heteròtrofa. Els consumidors s'alimenten d'altres éssers vius.
Poden ser:
- Primaris: herbívors, s'alimenten de vegetals - productors
- Secundaris: carnívors i s'alimenten d'herbívors. Alguns són omnívors: inclouen vegetals en la dieta.
- Terciaris: carnívors i s'alimenten d'herbívors i d'altres carnívors.
5.3 Descomponedors
Matèria orgànica morta — —► Descomponedors (fongs i bacteris) — —► Matèria inorgànica — —►Productors
6. Representacions de l'alimentació dels éssers vius en els ecosistemes
Piràmides tròfiques
Les relacions tròfiques són les relacions que s'estableixen entre els éssers vius que s'alimenten els uns dels altres.
Aquestes relacions es poden representar en forma de:
- Cadenes tròfiques: un seguit d'organismes que cada un és aliment del següent (és una cadena formada per baules interrelacionades entre elles)
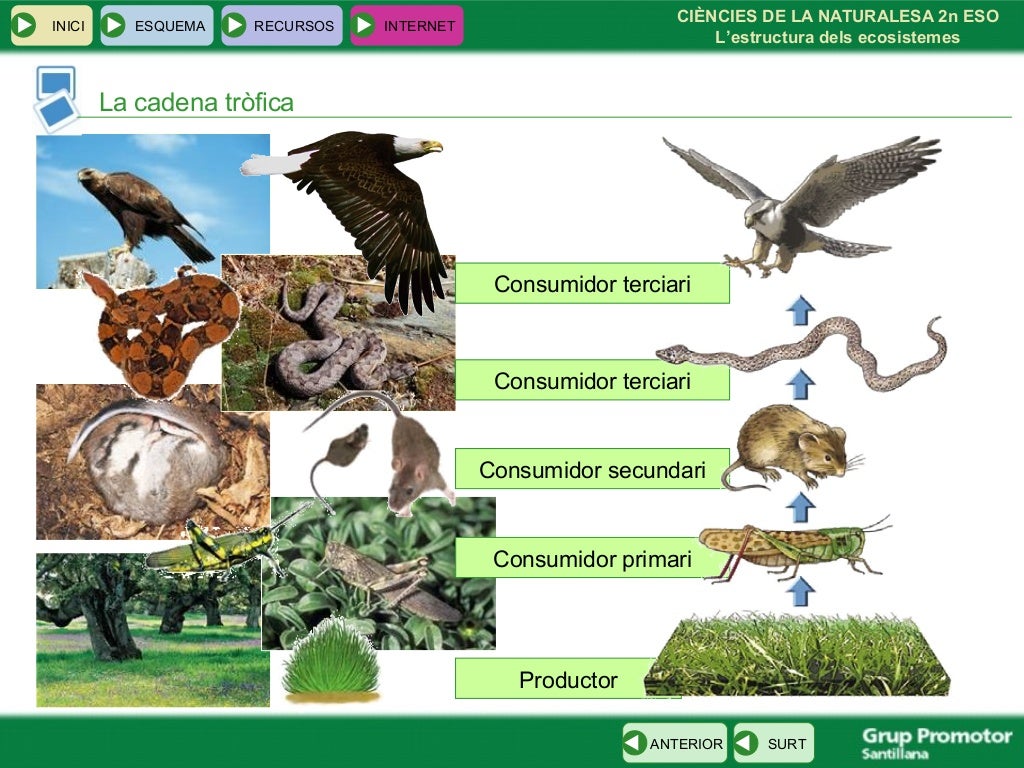
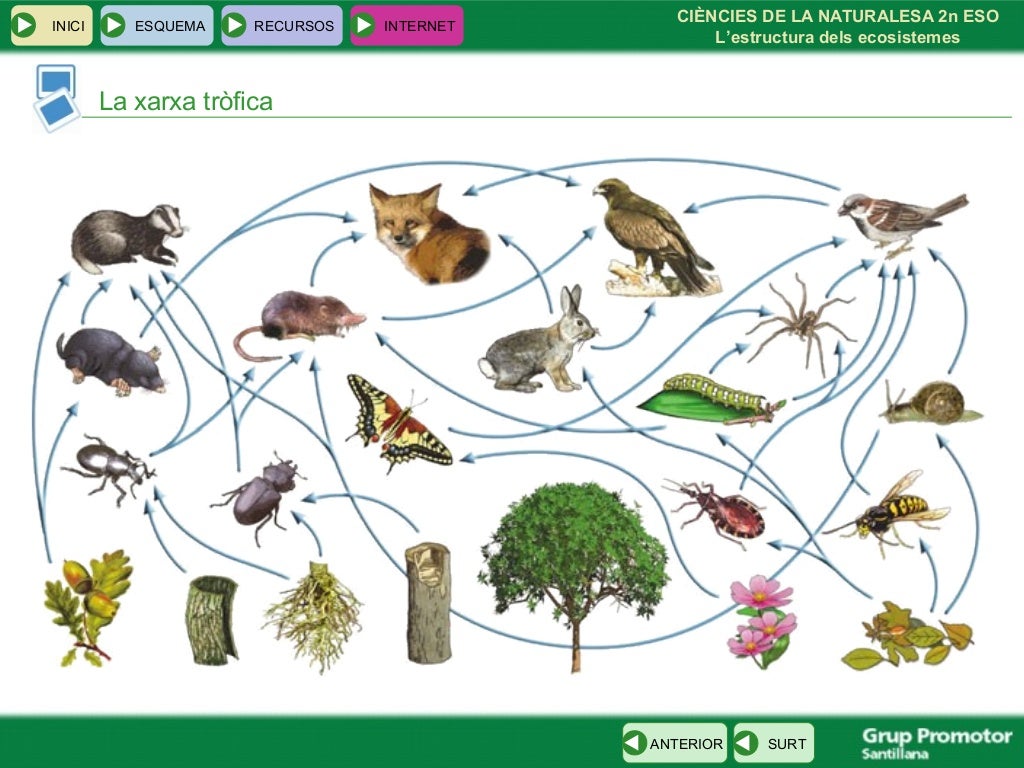
6.2 Piràmides tròfiques
Una piràmide tròfica és una manera de representar gràficament la variació que hi ha entre els diferents nivells tròfics segons una característica determinada (nombre d'individus, biomassa de cada nivell tròfic, energia de cada nivell tròfic).
6.2.1 Representació de les piràmides
- Les piràmides són pisos superposats.
- La base es correspon als organismes productors, el segon pis correspon als consumidors primaris, el tercer pis correspon als consumidors secundaris,...
- Tots els pisos tenen la mateixa alçada.
- L'amplada del pis és proporcional al valor que representa.

- No es té en compte la mida de l'organisme (un elefant i un llagost conten igual perquè són consumidors primaris).
- Cada pis representa la biomassa de cada nivell tròfic per unitat de superfície o per unitat de volum de l'ecosistema.
- Només poden ser normals.


7. La matèria i l'energia en els ecosistemes.
7.1 Flux d'energia
- L'energia entra a l'ecosistema com a energia lluminosa (Sol).
- Els organismes fotosintètics - autòtrofs transformen l'energia lluminosa en energia química (matèria orgànica)
- L'energia química dels productors és aprofitada ,en part, pels consumidors primaris. Part és perd en forma de calor en la respiració cel·lular.
- L'energia química dels consumidors primaris és aprofitada ,en part, pels consumidors secundaris. Part és perd en forma de calor en la respiració cel·lular.
- L'energia química dels consumidors secundaris és aprofitada ,en part, pels consumidors terciaris. Part és perd en forma de calor en la respiració cel·lular.
Aproximadament, el 10% de l'energia d'un nivell queda a disposició de l'altre nivell.
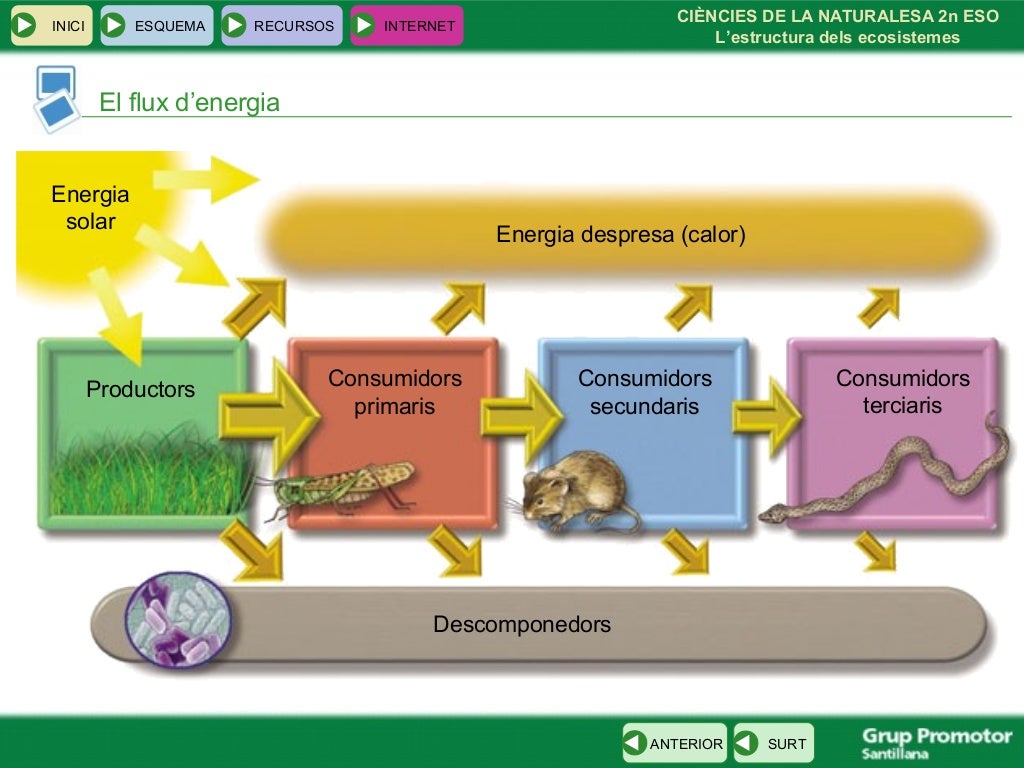
7.2 Cicle de la matèria
La matèria es recicla i no es perd. La matèria circula per l'ecosistema de forma cíclica.
- Els organismes productors fabriquen matèria orgànica a partir de matèria inorgànica.
- La matèria orgànica dels productors es consumida - menjada pels consumidors.
- La matèria orgànica dels consumidors primaris es consumida - menjada pels consumidors secundaris.
- La matèria orgànica dels consumidors secundaris es consumida - menjada pels consumidors terciaris.
- Quan es moren els productors i consumidors 1r, 2n, 3r els descomponedors transformen aquesta matèria orgànica morta en matèria inorgànica que torna a estar a disposició dels productors.
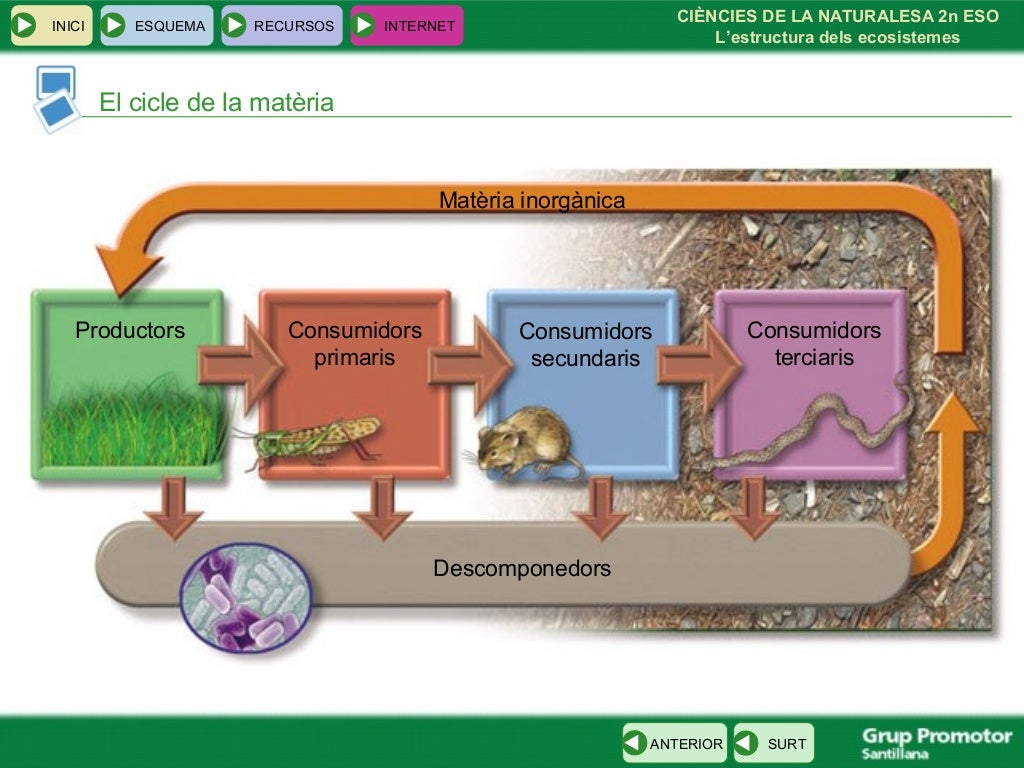
8. Relacions biòtiques. Relacions entre organismes vius.
Les relacions poden ser intra especígfiques o interespecífiques.
8.1 Relacions intraespecífiques
Tipus de relacions intraespecífiques:
famílies matriarcals lleones
famílies patriarcals elefants
Tipus de relacions interespecífiques:
Simbiosi - líquen.
- Comensalisme (+/0): un individu - comensal s'alimenta de les restes de mejar o productes alliberats per un altre organisme, al qual li és indiferent.
- Inquilinisme (+/0): un individu - inquilí es refugia en el cos o en algun resta d'un altre éssers viu sensen rejudicar-lo.
- Depredador - presa (+ depredador/ - presa): un individu - depredador mata i consumeix totalment o parcialment un altre individu - presa per alimentar-se.
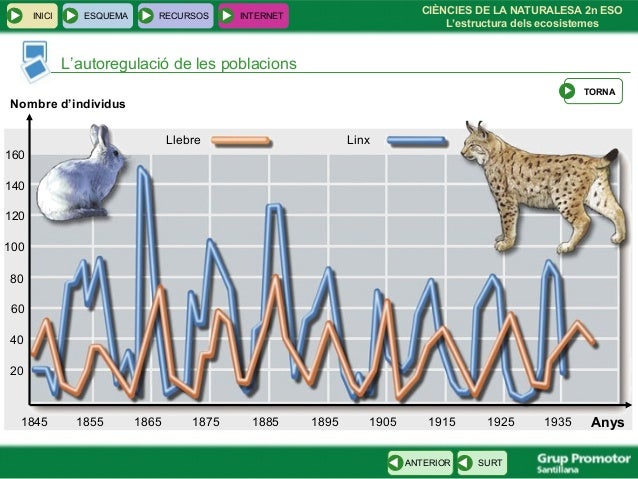
- Si representam en una gràfica el nombre d'individus d'una població en funció del temps, per un medi natural o sigui amb limitacions de recursos i d'espai, s'obtendria una corba en forma de S.
K - capacitat de càrrega: nombre màxim d'individus d'una població que un medi pot sostenir en unes condicions determinades (aliment, espai). Les poblacions que esmantenen pròximes a la seva K - capacitat de àrrega es diu que es troben en equilibri.
Exemple: el crac americà
- Imagina que una espècie nova - invasora (crac americà) arriba a un ecosistema (riu) on no havia existit mai.
- Aquesta espècie no tindrà limitacions d'espai ni d'aliment i creixerà exponencialment - J.
- El nombre d'individus de cranc americà serà molt gran.
- Quan tingui limitacions d'espai, d'aliment, sorgiexin competidors, o depredadors naturals el nombre d'individus de cranc tendirà a estabilitzar-se.
- Creixement en S. Haurà arribat a la seva K - capacitat de càrrega per aquell riu. (El crans pot tenir diferents K segons el medi - riu on es trobi).